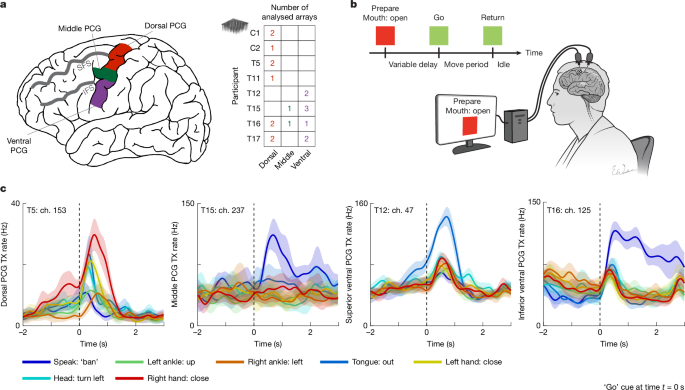
MainNearly a century ago, Penfield and colleagues pioneered the motor homunculus, a topographical map of the human PCG, by electrically stimulating the cortical surface and observing which body parts moved in response17,18. Since that time, the motor cortex has been mapped extensively in non-human primate models using single-neuron recordings6,7,8, microstimulation of both short and long durations1,2,3,4,5 and lesion and inactivation studies4,9,10. This body of work has provided strong evidence that the motor cortex is more fractionated and intermixed than originally implied by the motor homunculus, with related body parts overlapping in the primary motor cortex2,22,23 (for example, wrist and fingers) and whole-body regions overlapping in the premotor cortex6,7,24 (for example, arm and face). Long stimulation durations have more recently revealed that the motor cortex is also organized by behaviour in ethological ‘action maps’ as opposed to a pure body part organization3,4,21,25.In humans, however, exploration of the motor cortex has largely been limited to lower-resolution recording techniques—such as electrocorticography11,12,13 and functional magnetic resonance imaging (fMRI)14,15,16,26—as well as gross stimulation mapping in the operating room17,18,19,20. Studies reporting the effects of lesions27,28 and stimulation17,18,19,20 have found results that suggest a separation of leg, arm and face movement regions in the human PCG (that is, stimulation at any single location does not typically cause simultaneous arm, leg and face movement, with some exceptions19,29). On the other hand, fMRI and electrophysiological studies have emphasized a greater degree of intermixing13,14,15,16, including beta power modulation and blood-oxygenation-level-dependent deactivation spreading far from the somatotopic hotspot16,30, overlapping receptive fields14 and intereffector regions with whole-body intermixing15,31. However, these experiments cannot resolve detailed representations at the single-neuron level, and thus the representation of the body in the human motor cortex remains relatively unknown at a single-neuron scale. The crown (exposed surface) of the human PCG is thought to be anatomically composed largely of Brodmann area 6 (a premotor area), with the primary motor cortex lying within the central sulcus32,33,34. If the crown of the human PCG is functionally homologous to the macaque premotor cortex, we might expect to find dense intermixing of arm and leg movements dorsally3,7,24,35, and arm and orofacial movements ventrally3,6,24.In previous work using microelectrode arrays capable of recording brain activity at single-neuron resolution, we have shown that a small, anatomically distinct area of the cortex in the dorsal PCG (referred to as the ‘hand knob’36) contained intermixed representations of the entire body37,38 (including all four limbs and head and face movements), where the limbs were interrelated with a compositional neural code. However, it is unknown whether a similar organization exists in the middle and ventral PCG; in addition to providing fundamental insight into the neural representation of movement in humans, an answer to this question is needed to inform the design of brain–computer interfaces (BCIs) that restore motor function to people with paralysis, including speech.Here we revisit the motor representation of the whole body across a wide span of the PCG using microelectrode recordings from 8 human participants (Fig. 1a) implanted with 20 microelectrode arrays as part of BCI clinical trials. These participants had either spinal cord injury, amyotrophic lateral sclerosis (ALS) or brainstem stroke, resulting in severe motor impairment with varying levels of residual movement across individuals. All clinical trial procedures were conducted with oversight from ethics, institutional and regulatory bodies; before enrolment, an extensive consent process engaged potential participants and reinforced that no direct benefit from study participation was expected (see Methods). These neural recordings collectively sampled the length of the PCG spanning from the canonical arm area (near the superior frontal sulcus) to the canonical tongue and throat area (near the Sylvian fissure), and together constitute, to our knowledge, the first comprehensive motor map of the crown of the human PCG at single-neuron resolution.Fig. 1: Sampling the length of the PCG to assess whole-body movement representation.a, Diagram of the dorsal, middle and ventral regions of the PCG sampled in this study. Neural activity was analysed from a total of 20 microelectrode arrays distributed across 8 participants with paralysis (right table). Four additional microelectrode arrays were recorded but excluded from analysis due to lack of tuning to any movement (see Methods). All arrays were placed in the left PCG (and all participants except T12 and T15 were right handed). IFS, inferior frontal sulcus; SFS, superior frontal sulcus. b, Neural tuning to 46 attempted movements across the body was evaluated for each participant using an instructed delay task while they were positioned upright, either seated in a chair or in a bed at an incline. c, Example responses from microelectrodes within the three sampled regions of the PCG that show broad tuning to speech and movements of the arms, legs, face and head. Each line shows the mean threshold crossing (TX) rate across all trials of a single-movement condition, and the shaded regions show 95% confidence intervals (CIs). TX rates are a measure of spiking activity on an electrode and were denoised by convolving with a Gaussian smoothing kernel (120 ms s.d.). Black vertical dashed lines indicate the ‘go’ cue. Ch., channel.Neural tuning to the whole body in the PCGWe assessed neural tuning to speech and movements of the face, head, arms and legs in eight participants in a visually cued movement task (Fig. 1b and Supplementary Tables 1 and 2). If participants could not physically complete a movement due to paralysis, they were instructed to attempt to complete the movement in a light, controlled and isolated manner, to the best of their ability (see Supplementary Video 1 for example movements from T12, who retained the greatest residual arm and leg movement among the participants). Example electrodes demonstrating strong neural tuning to various movements are shown in Fig. 1c.To quantify neural tuning at a coarse scale, we measured the degree to which the neural population activity evoked by each movement differed from a ‘do nothing’ baseline condition (Fig. 2a). The majority of electrode arrays (15 of 20) recorded robust neural modulation in response to all tested movements, with the remainder showing modulation to almost all movements. We also analysed neural tuning at an individual electrode level and found that intermixed tuning to all body regions was frequently present within single electrodes (Extended Data Fig. 1) and spike-sorted single neurons (Extended Data Fig. 2).Fig. 2: Regional organization of whole-body movement tuning in the PCG.a, Neural modulation strength for each attempted movement was quantified by comparison to a ‘do nothing’ condition within each electrode array. The matrix is row-wise normalized to better visualize relative modulation strength (the brighter colours indicate larger differences between movement-related activity and rest activity). A white ‘X’ denotes non-significant modulation. In general, each array features broad tuning to all movements, with region-specific trends in tuning strength. Arrays are coded by participant identifier and region (dorsal (d), middle (m) and ventral (v)), with subscripts that order multiple arrays from the same participant in a medial-to-lateral sequence. All arrays were placed in the left hemisphere. b, Approximate spatial organization of arrays along the PCG relative to the anatomical landmarks of the SFS and IFS. Each array (square) is coloured in relation to its normalized modulation strength for a given movement class. Arm preference is a general feature near the SFS (canonical hand-knob area), whereas speech preference is seen near the inferior and middle regions of the PCG, flanking a more broadly tuned orofacial region near the IFS (canonical face-head area).Although all regions of the PCG were tuned to the whole body, sorting the arrays by anatomical location revealed that larger neuronal firing rate modulation aligned with the canonical motor homunculus, with arrays in the dorsal PCG showing a preference for arm movements, and arrays in the ventral PCG preferring orofacial movements (Fig. 2b). However, two findings also diverged from the classical homunculus. First, a majority of the arrays in the canonical orofacial region preferred speech to orofacial movement, particularly the most inferior ventral arrays (for example, T16-v1) and one middle PCG array (T15-m1). Second, the three arrays in between these speech-preferring arrays were more broadly tuned to the whole body. Arrays T12-v1 and T15-v1 in particular had substantial limb tuning and were also most strongly activated by orofacial movements as opposed to speech.Whole-body decoding from PCG sitesNext, to investigate the extent to which the whole body is represented in a movement-specific way throughout the PCG, we tested whether a recurrent neural network decoder could classify between movements belonging to the same category using neural activity from single trials (Fig. 3). With the exception of speech, where four similar words could not be classified above chance in dorsal arrays, classification accuracy within each movement category was above chance in all arrays (Fig. 3a and Supplementary Table 3). This indicates that a differentiable whole-body representation exists at each point in the human motor cortex, but with a different emphasis depending on the location (with the dorsal PCG showing greater decodability of limb movements, and the ventral PCG showing greater orofacial and speech decodability).Fig. 3: The PCG contains a decodable whole-body representation at each sampled location.a, Heatmap depicting classification accuracies from a recurrent neural network decoder trained to classify movements using single-trial neural recordings from each array separately. The red ‘X’ denotes classification accuracies not significantly above chance. With the exception of speech in dorsal arrays, movements belonging to the same category could be distinguished from each other above chance for all movement categories in all arrays. Of note, superior ventral arrays T15-v1 and T12-v1 express some of the highest decoding accuracies when classifying across all 45 movements (excluding the ‘do nothing’ condition). b, Example confusion matrix for the superior ventral array T12-v1, showing high accuracy across all movement types despite being a canonically orofacial area. RNN, recurrent neural network.Of note, the two most superior ventral arrays (T12-v1 and T15-v1) had some of the highest classification accuracies across all movement types. Figure 3b shows an example confusion matrix between all movements using activity from array T12-v1, which had an average 86% decoding accuracy. This result further supports the idea of a superior ventral orofacial zone with broad tuning to the whole body, but relatively weaker tuning to speech (than the speech zones in the middle PCG and inferior ventral PCG). Note that the neural representation of non-canonical movements (for example, orofacial movements in the dorsal PCG) was both multidimensional and relatively large (40% as large as that of canonical movements; Extended Data Fig. 3), suggesting that it may not simply reflect generic deactivation16 or minor spillover from nearby areas.When interpreting the classification results above, it is important to keep in mind three potential confounds: (1) some arrays simply record more tuned neurons than others, increasing their classification accuracy in general; (2) potential variability in how precisely individual movements are executed or attempted; and (3) some categories may be harder to classify than others due to the particular selection of movement conditions (for example, speech, in which the four tested words are very similar to each other). The third confound could explain why classification accuracy for speech is generally lower than other categories of movement, in contrast to the normalized firing-rate modulation results above, in which many arrays were speech dominant.Four functional zones in the PCGNext, we applied principal component analysis (PCA) to the normalized firing-rate modulation profiles and classification accuracies of each array to visualize array-tuning characteristics in a low-dimensional space (Fig. 4a). PCA identified two major axes that explained 73% of the variance in tuning profiles: a ‘speech–face versus limb’ axis that discriminated the inferior ventral arrays (and the middle PCG array T15-m1) from the dorsal arrays, and a ‘breadth of tuning’ axis that discriminated the superior ventral arrays from the others. The coefficients for these two PCA axes (Fig. 4b) were similar for both the firing-rate modulation profiles (solid lines) and the classification accuracies (dashed lines), indicating general agreement between these two complementary methods of assessing whole-body tuning.Fig. 4: Four functional zones in the PCG.a, Low-dimensional representation of the tuning properties of each array, generated by applying PCA to the neural-tuning magnitudes and classification accuracies associated with each array. Arrays are coloured according to their position in the PCA space (the colours are used to construct the topographical map of tuning in panel c). b, Coefficients of the top two principal components. The first principal component (PC1) describes a speech–face versus limb axis, whereas PC2 describes the breadth of tuning. The solid lines denote coefficients related to modulation strength, and the dashed lines denote coefficients related to classification accuracies. c, Arrays are shown topographically and coloured according to their position in the PCA space (the colours match panel a), revealing functional zones of tuning properties. Combining this information with what is known in the literature, we hypothesize that there are four functional zones in the sampled region of the PCG: inferior ventral and middle zones related to speech (orange), a superior ventral orofacial area that also contains strong arm–hand tuning (purple), and one arm–hand dominant zone (cyan).We coloured each array according to its location along the top two PCA axes, and displayed this information topographically to reveal regional patterns in tuning characteristics (Fig. 4c). On the basis of these results, as well as what is known from previous work (particularly studies identifying the middle PCG as a speech–language region32,39), we hypothesize that there are four main functional zones in the sampled area of the PCG (Fig. 4c): one dorsal zone emphasizing arm movements, one inferior ventral zone emphasizing speech, one middle PCG zone also emphasizing speech (array T15-m1), and one superior ventral orofacial zone between the speech zones that is most broadly tuned to the whole body (arrays T12-v1 and T15-v1). We also analysed fMRI resting state data from the Human Connectome Project32 and found evidence of two resting-state networks that parcellate the PCG into four zones (Extended Data Fig. 4): one language-related resting-state network with hotspots in the inferior ventral PCG and middle PCG (accounting for the speech zones), and one hand–arm-related resting-state network with hotspots in the dorsal PCG and superior ventral PCG (which could explain the strong limb tuning in the superior ventral PCG).It is possible that anatomical variability across individuals could cause other zones to sometimes appear on the crown of the PCG. Array T16-m1 in particular appears to be an outlier in exhibiting strong head-related modulation and classification accuracy. Using the Human Connectome Project cortical parcellation procedure applied to the preoperative neuroimaging data for T16, we identified array T16-m1 as being on the border of the premotor eye field, an area that may be highly tuned to movements of the eyes and head and that typically lies within the precentral sulcus (Extended Data Fig. 5).Finally, using preoperative neuroimaging data obtained from four participants (T12, T15, T16 and T17), we were able to assess array locations relative to recently identified inter-effector regions in the somatocognitive action network15, as well as the hand–face border40 (Extended Data Fig. 6). We found that all arrays appeared to lie outside of the inter-effector regions. In addition, the superior ventral arrays with broad whole-body tuning (T12-v1 and T15-v1) were placed below the hand–face border and within the canonical face region of the motor cortex. These results show that strong whole-body tuning is not limited to previously identified inter-effector regions or to the hand–face border.A compositional whole-body neural codeFinally, we sought to elucidate the representational structure of the whole-body code found throughout the PCG. In previous work investigating the hand-knob area37, we found a ‘compositional’ neural code linking all four limbs together that had two main features: (1) correlated representations of homologous movements across limbs (for example, wrist flexion and ankle flexion), and (2) representation of the limb itself independent of the movement.Here we also found strong correlations between all four limbs in multiple regions of the PCG. Figure 5a shows example arm–leg correlations for the dorsal PCG and superior ventral PCG (Extended Data Fig. 7 shows the correlation matrices for all regions and limb pairs). Representational similarity between limbs was present for most limb pairs and most arrays (Fig. 5b), although the arms appeared to be most strongly correlated with one another (significant for 17 of 20 arrays). These results show that inter-limb neural correlations linking all four limbs are a general feature of the human motor cortex, appearing even in orofacial and speech regions. Using video data from participant T12, who could still make visible arm and leg movements, we confirmed that this effect is not the result of inadvertent movement of uninstructed effectors (Extended Data Fig. 8 and Supplementary Video 1).Fig. 5: A compositional neural code throughout the PCG that links all four limbs together.a, Representational similarity of neural activity for homologous arm and leg movements in two example regions of the PCG. Each square in the matrix represents pairwise similarity for two movements (as measured by the correlation between neural firing-rate vectors). Ordering movements according to their arm–leg homologue shows a diagonal structure indicating high similarity between homologous movements. b, Summary of representational similarity for each electrode array across various limb pairs. Each square in the matrix represents the average similarity between homologous movements of a limb pair (as measured by the mean of the diagonal values of the corresponding similarity matrix). A black ‘X’ indicates non-significant similarity via shuffle testing. c, Low-dimensional PCA visualizations of single-trial neural activity for four example arrays (each trial is a single circle that is coloured according to the side of the body; only arm movement conditions are plotted for clarity). The dorsal arrays show a clear ‘laterality dimension’ that separates the right and left arms, whereas the middle and ventral arrays feature a more intermixed representation in the top two principal components. d, Summary of the size of the laterality-related variance for arm movements relative to other factors of the neural data. Data were marginalized according to the following factors: time, laterality, movement and the laterality–movement interaction. Generally, the laterality dimension was stronger for the dorsal arrays than for the middle and ventral arrays (as measured by the fraction of variance within the marginalization).Next, we tested for the existence of limb-coding activity in the form of ‘laterality’ neural dimensions that would separate the left from right arms independently of the movement itself, as found previously in the hand-knob area37,41. Consistent with previous results, we found that a laterality dimension is a dominant feature of the dorsal PCG, appearing clearly in the top two principal components of the neural activity (Fig. 5c; Extended Data Fig. 9 shows scatterplots for all arrays). Laterality information was less prevalent in other areas of the PCG (Fig. 5c,d), indicating that this feature is probably unique to the dorsal PCG.DiscussionHere we created a comprehensive motor map of the crown of the human PCG at single-neuron resolution, leveraging microelectrode array recordings from 20 arrays across 8 individuals with paralysis (Extended Data Fig. 10). This map revealed that distinguishable, multi-dimensional whole-body representations exist at all sampled points of the PCG. We also found that these whole-body representations are abstract, linking together the legs and arms in a partially limb-independent representation of motor action. A limb-independent movement code could facilitate the transfer of learned actions from one limb to another42,43, and was present throughout all sampled areas of the PCG. Our results also shed new light on the regional arrangement of the human motor cortex. We found evidence that the sampled extent of the PCG between the Sylvian fissure and superior frontal sulcus appears to contain four functional zones, each with their own whole-body representations: (1) a dorsal arm–hand zone, (2) a middle speech zone (also highlighted in recent work32,39), (3) a broadly tuned, superior ventral orofacial zone, and (4) an inferior ventral speech zone.Our results support a growing body of evidence that the motor cortex is more integrative than implied by the classical homunculus. Recent results from lower-resolution recording methods in humans (for example, fMRI and electrocorticography) have demonstrated overlapping body part representations in the PCG13,14,15,31. In animal models, the idea of ethological action maps as an organizational principle for the motor cortex has come to prominence in the past few decades3,4,21,25. Long-train stimulation delivered to premotor areas creates complex, multi-joint action patterns that can span body regions and appear to be pieces of ethological behaviours3. This theory could explain the overlapping whole-body representations that we observed in the PCG. For instance, the dorsal PCG could implement behaviours that coordinate the limbs, whereas the ventral PCG may implement behaviours involving combined orofacial and limb movement (for example, self-feeding); more broadly, the entire body may be involved in many behaviours due to the need to stabilize the limbs, trunk and head. Our findings of speech-specific areas in the PCG also support the idea of a behavioural organization in the motor cortex.Although the crown of the human PCG is sometimes referred to as the primary motor cortex19,20, anatomical evidence has shown that the crown of the PCG in humans is composed largely of Brodmann area 6 (a premotor area), whereas Brodmann area 4 (primary motor cortex) lies within the central sulcus32,33,34. Our results support a homology between the crown of the human PCG and the macaque premotor cortex, which is known to contain more intermixed6,7,24 and abstract44,45 movement representations than the primary motor cortex. Specifically, the superior ventral region of the PCG was strongly tuned to both orofacial and arm–hand movement, and may be homologous to the macaque ventral premotor cortex, which is known to contain grasping, reaching and orofacial-related information6,46,47 (see also ref. 48); likewise, the dorsal PCG may be homologous to the macaque dorsal premotor cortex, which contains both forelimb and hindlimb movement representations5,7,24,35,47. A recent cortical atlas based on neuroimaging data from the Human Connectome Project32 has also divided the crown of the human PCG into a series of premotor areas that are distinct from the central sulcus (Extended Data Fig. 5).The degree of intermixing observed here appears to be at odds with electrical stimulation studies in people, which generally have revealed separate regions for leg, arm and face movements that do not often intermix at the same location, consistent with Penfield’s original homuncular diagram (despite some exceptions found in one-fifth of patients19). Lesions to the human motor cortex have also been reported to cause focal deficits49,50 (for example, lesions to the hand-knob area have been reported to affect only arm and hand function). This could indicate that the whole-body representations observed here, although relatively large (approximately 40% the size of canonical movement representations), may be ancillary to the main function of the areas, for instance, reflecting inputs from neighbouring regions that support coordination of movements involving multiple effectors. However, other interpretations are possible. For instance, it may simply be the case that electrical stimulation activates mainly the strongest representation (especially when stimulating for short durations). In addition, lesions are not often contained only in the grey matter on the crown of the PCG, and may cause focal deficits due to white matter damage or damage to adjacent primary motor areas in the central sulcus.One important question is whether the results shown here would generalize to able-bodied individuals without spinal cord injury, brainstem stroke or ALS. The dense intermixing of the whole body found at each sampled point and within each participant, as well as the limb-independent movement code found across all participants, suggest that these features are not specific to a particular disease or level of paralysis (see Supplementary Table 4 for motor strength scores). Although our results from the ventral PCG are limited only to individuals with brainstem stroke or ALS, results still reproduced for different stages and levels of paralysis (for example, T15 and T12 both showed broad orofacial and limb tuning in the superior ventral PCG, despite T15 being much more paralysed than T12). In addition, a large body of work has shown that the extent of motor reorganization in the adult brain after amputation is quite limited, suggesting that loss of limb function does not greatly alter motor topography51,52 (see also recent neuroimaging results in spinal cord injury53). Furthermore, although myelin and motor cortical function is known to degrade in ALS54,55, it is not clear why this would cause body parts to intermix more than they otherwise would normally. Finally, it is possible that people with severe motor impairment may unintentionally co-activate other effectors to compensate for their weakness when attempting isolated movements, potentially recruiting neural representations of other body parts. We minimized this possibility by coaching participants to attempt movements in a light, controlled manner and confirmed in T12—who retained visible arm and leg movements—that tuning to all four limbs was not due to overt co-movement. However, most other participants had more limited residual movement, so comparable behavioural verification was not feasible to rule out subtle co-activation. Nonetheless, only recordings from able-bodied people can provide a definitive answer as to whether these results generalize to healthy individuals.Finally, our study charts the first map of how well movements from each category can be decoded at single-neuron resolution throughout the PCG. This map can inform the design of intracortical BCIs aiming to restore speech, arm, hand and leg movement to people with paralysis56,57,58. To achieve the highest performance with the minimal number of electrodes, our results show that speech BCIs may benefit from targeting the inferior ventral and middle PCG while avoiding the superior ventral region. By contrast, the superior ventral region may be an underappreciated complement to the dorsal PCG for arm–hand BCIs48,59. Overall, the widespread decodability of the whole body throughout the PCG is advantageous for BCIs seeking to restore multiple functions with a small footprint, although not all body parts are equally decodable from all locations60.MethodsExperimental proceduresStudy participantsThis study includes data from eight participants who each gave informed consent before any experimental procedures. Participants T5, T11, T12, T15, T16 and T17 were enrolled in the BrainGate2 Neural Interface System clinical trial (ClinicalTrials.gov Identifier: NCT00912041, registered 3 June 2009), and represent all BrainGate2 participants who were enrolled at the time the data were collected and analysed. This pilot clinical trial was approved under an Investigational Device Exemption (IDE) by the US Food and Drug Administration (FDA; Investigational Device Exemption #G090003). Permission was also granted by the Stanford University Institutional Review Board (IRB; protocol #20804), the Mass General Brigham IRB (protocol #2009P000505), the University of California, Davis IRB (protocol #1843264), the Emory University IRB (protocol #00003070) and the Providence VA Healthcare IRB. Participants C1 and C2 were enrolled under a separate multi-site clinical trial (ClinicalTrials.gov Identifier: NCT01894802, registered 10 July 2013), which was also conducted under an IDE from the US FDA and approved by the IRBs at the University of Pittsburgh and the University of Chicago. C1 and C2 represent all participants enrolled under NCT01894802 who were available at the time of data collection to participate in this study, given competing demands on participant time for other research goals. All research was performed in accordance with relevant guidelines and regulations.Recognizing that cortical mapping and neural activity recording in humans introduces distinctive ethical and philosophical considerations, ethics oversight was embedded from the outset. All risks (including surgical risks) arise solely from the parent safety and feasibility trials (and not the present study), each approved and monitored by local IRBs. Additional safeguards were provided by multiple and in part redundant additional layers of protection, including continuous oversight from local Clinical Oversight Committees including an independent medical monitor, a Data Safety and Monitoring Board, strict adherence to FDA guidance on conduct and safety under our IDE, clinical trial offices from the US National Institutes of Health (NIH), and engagement with the Neuroethics Working Group (NEWG) of the NIH BRAIN Initiative, to ensure adherence to the highest standards of safety and ethical rigour. All study procedures complied with the Declaration of Helsinki, the Belmont Report, CIOMS guidance and the NIH BRAIN Neuroethics Principles. In the parent clinical trials, a months-long, dialogue-driven consent process safeguards participant autonomy and adapts to evolving communication needs, protecting participant privacy. During this process, we explained that there would be no direct benefits from the implanted device. Within the trials, proactively embedded neuroethics expertise encourages continual reflection on risk–benefit balance, participant welfare and societal impact.T5 is a right-handed man, 69 years of age at the time of this study, with tetraplegia due to cervical spinal cord injury (classified as C4 AIS-C), which occurred approximately 9 years before enrolment in the clinical trial. T5 has two 96-channel intracortical microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the hand-knob area of the left PCG. The hand-knob area was identified anatomically by preoperative MRI. T5 has full movement of the face and head and the ability to shrug his shoulders. Below the level of the spinal cord injury, T5 has very limited voluntary motion of the legs and arms.T11 is a right-handed man, 38 years of age at the time of this study, with tetraplegia due to a cervical spinal cord injury (classified as C4 AIS-B), which occurred approximately 14 years before. T11 has two 96-channel intracortical microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the left dorsal PCG, targeting the hand-knob area as identified anatomically by preoperative MRI. T11 has full movement of the face and head with very limited voluntary motion of the arms.T12 is a left-handed woman, 67 years of age at the time of this study, with slowly progressive bulbar-onset ALS. T12 was diagnosed at age 59 with an ALS-Functional Rating Scale score of 26 at the time of study enrolment. T12 has four 64-channel intracortical microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the left (language dominant, motor non-dominant) hemisphere, based on preoperative anatomical MRI, fMRI and cortical parcellation using the Human Connectome Project (HCP)32 multi-modal parcellation pipeline (see Extended Data Fig. 5 for cortical parcellation results). Two arrays were placed in the inferior frontal gyrus (not included in this study), and two arrays were placed in the ventral PCG, targeting area 6v; see Willett et al. for more details56. T12 remains functionally independent with 3–4 out of 5 strength (assessed using the Medical Research Council (MRC) scale) in all limbs, but is anarthric (able to vocalize, but unable to produce intelligible speech).T15 is a left-handed man, 45 years of age at the time of this study, with ALS (ALS-Functional Rating Scale score of 23 at the time of study enrolment). T15 has four 64-channel intracortical microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the left (language dominant, motor non-dominant) PCG, based on preoperative anatomical MRI and HCP cortical parcellation (see Extended Data Fig. 5 for cortical parcellation results). One array was placed targeting area 55b, two arrays targeting area 6v and one array targeting area 4; see Card et al. for more details61. T15 has limited orofacial movement with the capacity for vocalization, but is unable to produce intelligible speech. T15 has very limited voluntary motion of the rest of the body.T16 is a right-handed woman, 52 years of age at the time of this study, with tetraplegia and dysarthria due to a pontine stroke approximately 19 years before enrolment in the BrainGate2 clinical trial. T16 has four 64-channel intracortical microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in her left PCG: two in the hand-knob area (targeting area 6d), one in the speech-related ventral premotor cortex (targeting ventral area 6v) and one in the middle PCG (targeting area 55b). Implant area targets were identified by the HCP multimodal cortical parcellation procedure (see Extended Data Fig. 5 for the cortical parcellation results). Examination of post-implant array locations indicated that the middle PCG array appears to be on the border between area PEF and 55b. T16 is able to speak slowly and quietly, but speech cadence is reduced due to poor diaphragm voluntary control. She has limited voluntary control of her upper extremities, with some shoulder motion and some slow and contractured wrist and finger movements. She has limited-to-no voluntary control of her lower extremities. Sensation for T16 is fully intact.T17 is a right-handed man, 33 years of age with a history of rapidly progressive ALS. T17 has six 64-channel intracortical microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the left hemisphere, based on preoperative anatomical MRI, task-based fMRI and cortical parcellation using the HCP multimodal parcellation pipeline. Two arrays were placed in the dorsal PCG (targeting area 6d), two arrays were placed in the ventral PCG (targeting area 6v) and two arrays were placed in area 55b. At the time of this study, T17 is quadriplegic, anarthric and ventilator dependent. His only remaining volitional motor control is over his extra-ocular movements.C1 is a right-handed man, 57 years of age at the time of implant, who presented with a C4-level ASIA-D spinal cord injury that occurred 35 years before implant. C1 has four microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the left hemisphere. Two 96-channel microelectrode arrays were implanted in the arm and hand area of the motor cortex and two other 32-channel arrays (not included in this study) were implanted in the somatosensory cortex. Targeted array placement was based on functional neuroimaging (fMRI) of the participant attempting to make movements of the hand and arm, within the constraints of anatomical features such as blood vessels and cortical topography; see Greenspon et al. for more details62. C1 had no control of the intrinsic or extrinsic muscles of the right hand but retained the ability to move his arm with noted weakness in many upper limb muscles. He retained impaired, but largely functional, movement of the other limbs, full control of head and face movement, and could speak fluently. The data included here were collected 3.25 years post-implant.C2 is a right-handed man, 60 years of age at the time of implant, who presented with a C4-level ASIA-D spinal cord injury and right brachial plexus injury that occurred 4 years before implant. C2 has four microelectrode arrays (Blackrock Microsystems; 1.5-mm electrode length) placed in the left hemisphere. Two 96-channel microelectrode arrays were implanted in the arm and hand area of the motor cortex and two other 32-channel arrays (not included in this study) were implanted in the somatosensory cortex. Targeted array placement was based on functional neuroimaging (fMRI and magnetoencephalography) of the participant attempting to make movements of the hand and arm, within the constraints of anatomical features such as blood vessels and cortical topography62. C2 retained full control of his entire body except for right hand and arm movement. He could speak fluently. The data included here were collected 0.75 years post-implant.Neural signal processingFor each participant, neural signals were recorded from the microelectrode arrays using the NeuroPort system (Blackrock Microsystems). The signals were then analogue filtered (fourth-order Butterworth with corners at 0.3 Hz to 7.5 kHz) and digitized at 30 kHz (250-nV resolution). The subsequent digital filtering and neural feature extraction methods differed between participants due to variations in the systems at different sites.For T5, T11 and T12 the signals were decimated to 15 kHz and band-pass filtered between 250 Hz and 4,900 Hz using a fourth-order zero-phase non-causal Butterworth filter. Linear regression referencing (LRR) was then applied to further reduce ambient noise artefacts63 before spike detection. Spike threshold crossing detection was implemented using a −4.5 × RMS threshold applied to each electrode, where RMS is the electrode-specific root mean square of the time series voltage recorded on that electrode.For T15, the signals were band-pass filtered between 250 Hz and 5 kHz using a fourth-order zero-phase non-causal Butterworth filter, and LRR was then used to reduce noise artefacts. Spike threshold crossing detection was implemented using a −4.5 × RMS threshold.For T16, each electrode was high-pass filtered with a 250 Hz cut-off using a fourth-order zero-phase non-causal Butterworth filter. LRR was used for noise reduction and artefact removal with parameters computed post-filtering from a dedicated reference block at the beginning of the session. A −3.5 × RMS threshold was applied to each electrode for spike threshold crossing detection.For T17, the signals were decimated to 15 kHz and band-pass filtered between 250 Hz and 5,000 Hz using a fourth-order zero-phase non-causal Butterworth filter. LRR was then applied to further reduce ambient noise artefacts before spike detection. Spike threshold crossing detection was implemented using a −3.5 × RMS threshold applied to each electrode.For participants C1 and C2, a high-pass filter (250 Hz) was applied to each electrode before spike detection. Spike threshold crossing detection was implemented using a −4.5 × RMS threshold applied to each electrode.The resulting spiking data from each participant mentioned above was binned in 20-ms bins for offline analyses and decoding as presented throughout this study.Overview of data collection sessions and cued movement taskFor each participant, neural data were recorded in a single ‘session’ on a scheduled day. During the session, the participant was seated in a chair in front of a computer monitor at an idle and relaxed position (with the exception of participants T5 and T17, who were lying in a bed at an incline). Each participant completed a series of 5–10 min ‘blocks’ of the cued movement task, consisting of an uninterrupted series of trials. Supplementary Table 1 lists all data collection sessions reported in this work. Variation in the number of trials and/or blocks collected for each participant is due to differences in session durations for each participant and their respective comfort and/or fatigue levels.The cued movement task followed a simple instructed delay paradigm (Fig. 1b). During the instructed delay period, a red square and text appeared in the centre of the screen indicating to the participant that they should prepare to make the specified movement. The instructed delay period varied randomly (except for participants C1 and C2), with the range tailored to preference of each participant for sufficient time to read the prompt and prepare the movement. After the delay, the square turned green and the text indicating the movement changed to ‘go’, at which point the participant executed the movement immediately. The participant was instructed to make the movement if they were able to overtly move that body part, otherwise they were instructed to attempt the movement. They were then directed to continue attempting the movement, or holding the posture of the completed movement until the text changed to ‘return’, at which point the participant relaxed and returned to a neutral posture. Typically, the movement period lasted 1.5 s and the return period lasted 1 s, although these durations were adjusted if participants required more time to complete the task. See Supplementary Table 2 for task timing parameters.A standardized document describing task details of the whole-body movement sweep was shared with each site before data collection. Participants were instructed to prioritize isolated and consistent attempted movements. The following text summarizes the key task elements and instructions provided in that document.The session design details included: “This session is to collect open-loop attempted movements for various effectors across the entire body. We ask the participant to attempt or make a single isolated movement which is cued via text on a computer monitor in a delayed movement task. As the participant attempts the cued movement, neural data is recorded”.Key notes included:
A mosaic of whole-body representations on the human precentral gyrus - Nature
A comprehensive map of the human motor cortex at single-neuron resolution is described.
15,824 words~72 min read