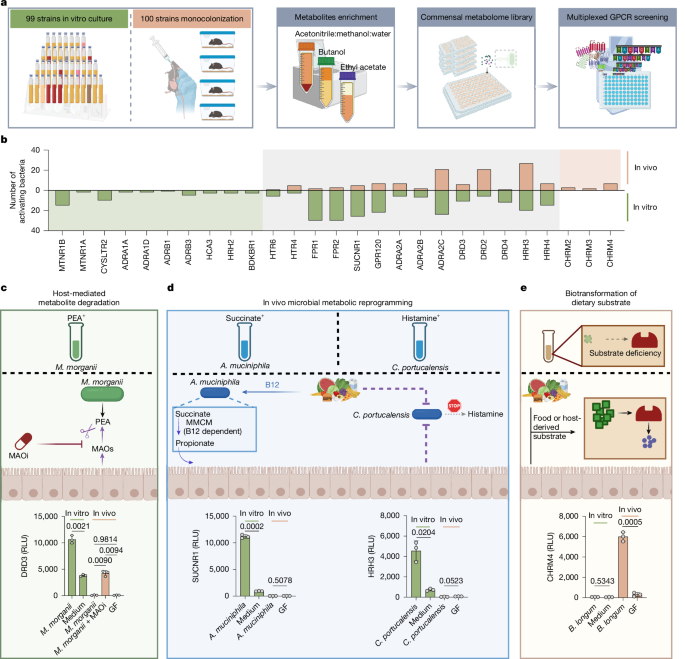
MainThe human gut microbiota produces thousands of small-molecule metabolites through de novo synthesis and through the biotransformation of chemicals derived from our diet or produced and secreted by the host8,9,10. These microbiota metabolites have essential roles in regulating diverse physiological functions, including energy metabolism, inflammation and mucosal barrier integrity11,12,13. Previous studies from our laboratory and others have used high-throughput GPCR screens of supernatants from commensal strains grown in vitro (that is, in vitro commensal metabolomes) to identify impacts of microbiota metabolites on host physiology4,6. However, the chemical diversity and physiological relevance of these in vitro metabolomes are constrained by the simplicity of the cultivation medium and the unavoidable artificiality of in vitro growth conditions. By contrast, in vivo microbiota metabolomes reflect the dynamic environment of the mammalian gastrointestinal tract, in which commensal microorganisms are exposed to complex environmental cues and precursor chemicals from the host, diet and other microorganisms. However, due to the requirement for large sample volumes, it is not possible to perform unbiased receptorome-wide GPCR screening of in vivo microbiota metabolomes using conventional approaches. To overcome this limitation, we recently developed a highly multiplexed GPCR screening platform (PRESTO-Salsa) that enables receptorome-wide assessment of nearly all non-olfactory GPCRs (>300 receptors) in a single well of a 96-well plate5 and used this technology to directly compare bioactivities of in vitro and in vivo commensal metabolomes.In vitro and in vivo metabolome screensThe microbial compositions of commensal communities grown in vitro differ substantially from those grown in vivo, making it infeasible to compare complex microbial communities grown under these two conditions. Thus, to enable direct comparisons between in vitro and in vivo metabolomes, we compared metabolomes from individual strains grown either in standard culture medium or in monocolonized gnotobiotic mice. We selected 100 phylogenetically diverse microbiota strains from public and internal strain collections (Supplementary Table 1). For in vitro bacterial metabolome libraries, we cultured each bacterial strain in medium and extracted using three different solvents known to enrich for distinct metabolite classes. For in vivo metabolomes, germ-free (GF) C57BL/6 mice were monoassociated with individual strains for 2 weeks, after which caecal contents were collected and extracted using the same solvents. We then screened the resulting 597 enriched commensal metabolomes (297 from in vitro cultures and 300 from mono-associated mice) against nearly all conventional GPCRs using PRESTO-Salsa (Fig. 1a and Supplementary Table 2).Fig. 1: In vivo and in vitro commensal metabolomes exhibit distinct GPCR activation patterns.The alternative text for this image may have been generated using AI.Full size imagea, Schematic of generating commensal metabolomes from bacteria cultured in vitro in medium and in vivo in monoassociated GF mice, followed by GPCRome interaction screening using the multiplexed GPCR screening technology PRESTO-Salsa. b, The number of in vitro or in vivo commensal metabolomes activating each GPCR. c–e, Representative models of in vivo-mediated alterations in GPCR-active commensal metabolites. c, Host MAOs degrade bacterially derived PEA in vivo. MAOi, MAO inhibitor; RLU, relative luciferase units. d, In vivo metabolic reprogramming suppresses succinate or histamine production. e, Diet-derived choline enables bacterial ACh production in vivo. In c–e, GPCR activity data from in vitro samples are representative of three independent experiments (n = 3 biologically independent samples per group in each experiment); GPCR activity data from in vivo samples are representative of two independent experiments (n = 3 mice per group in each experiment). For c–e, data are mean ± s.e.m. Statistical significance was assessed using two-sided Welch’s t-tests (d and e and in vitro comparison in c) and Welch’s analysis of variance (ANOVA) followed by Games–Howell post hoc test (in vivo comparison in c). The diagrams in a and c–e were created using BioRender; Song. D https://BioRender.com/x0hrt5l (a) and https://BioRender.com/sqnbzth (c–e) (2026).Source dataConsistent with our previous findings, select in vitro metabolomes activated adrenergic receptors (ARs), dopamine receptors (DRDs), histamine receptors (HRHs), serotonin receptors (HTRs), formyl peptide receptors (FPRs) and the succinate receptor4,5 (SUCNR1; Fig. 1b). Extraction-based enrichments also revealed activities at additional receptors that were not detected previously with crude supernatants, including the melatonin receptors (MTNR1A and MTNR1B), cysteinyl leukotriene receptor 2 (CYSLTR2), hydroxycarboxylic acid receptor 3 (HCA3), bradykinin receptor B1 (BDKBR1) and GPR120 (Fig. 1b and Supplementary Table 3).Compared with in vitro metabolomes, corresponding in vivo-derived commensal metabolomes exhibited distinct GPCR activation patterns. Among the GPCRs activated by in vitro metabolomes, most were also activated by in vivo metabolomes, including DRDs, ARs, HRHs, FPRs, HTRs, SUCNR1 and GPR120 (Fig. 1b), although the in vivo metabolomes of fewer strains exhibited these activities. Notably, select strains exhibited unique receptor engagements in vivo but not in vitro, particularly for the ARs α2A (also known as ADRA2A), ADRA2C, DRD2, HRH3 and HTR4 (Fig. 1b, Extended Data Fig. 1a,b and Supplementary Table 3). We created a publicly accessible interactive website (https://nwplab.shinyapps.io/commensal-vitro-vivo-metabolome-gpcr/) that highlights these differences and enables exploration of all screening data.Overall, the major differences in GPCR activation patterns between in vitro and in vivo metabolomes fell into at least three non-mutually-exclusive potential mechanistic categories (additional details and examples are provided in the Supplementary Note). First, host enzymes can degrade lumenal microbiota-derived metabolites, limiting their availability in vivo. For example, Morganella morganii NP17-derived phenethylamine (PEA) activated DRDs and ARs in vitro but not in vivo unless host monoamine oxidases (MAOs) were inhibited (Fig. 1c and Extended Data Fig. 2). Second, in vivo environments can induce reprogramming of microbial metabolism. For example, SUCNR1 activation was frequently observed in vitro but often diminished in vivo, in part due to dietary vitamin-B12-dependent metabolic rewiring in taxa such as Akkermansia muciniphila (Fig. 1d and Extended Data Fig. 3). A similar in vitro-to-in vivo shift was observed for histamine-related activities. Some commensals activated HRHs in vitro but not in vivo (Fig. 1d and Extended Data Fig. 4), and transcriptional analysis of Citrobacter portucalensis NWP28 revealed reduced expression of genes involved in amino acid decarboxylation and histidine uptake after gut colonization (Extended Data Fig. 5). Finally, biotransformations of diet- or host-derived substrates that are present in vivo and absent from standard culture medium can lead to the emergence of unique in vivo bioactivities. For example, multiple in vivo commensal metabolomes, but not their corresponding in vitro metabolomes, activated the muscarinic ACh receptors (CHRMs) (Fig. 1b,e), suggesting that microbiota strains may produce CHRM ligands by biotransforming host- or diet-derived precursors.Select commensals produce ACh in vivoOur GPCRome-wide screening studies revealed that in vivo metabolomes from multiple bacterial strains activated the CHRMs (as represented by CHRM4), including Bifidobacterium longum NWP308, Bifidobacterium adolescentis NWP330, Bacteroides caccae NWP468, Bacteroides fragilis NWP229, Bacteroides dorei NWP244, Ruminococcus gnavus ATCC29149, P. pentosaceus NP38 and Streptococcus parasanguinis NWP114 (Fig. 2a). Notably, none of these strains activated CHRM4 when cultured in vitro (Fig. 2a).Fig. 2: Select gut commensal strains produce ACh.The alternative text for this image may have been generated using AI.Full size imagea, CHRM4 activation (indicated by log2-transformed fold change (log2[FC])) by in vitro (bottom) and in vivo (top) commensal metabolomes as measured using PRESTO-Salsa. n = 3 biologically independent samples. b, CHRM4 activation by in vitro versus in vivo metabolomes from representative bacteria measured using PRESTO-Tango. c, Mass spectrometry quantification of ACh in the bacterial metabolomes analysed in b. d, CHRM4 activation by representative bacteria cultured in vitro with 1 mM choline. e, Mass spectrometry quantification of ACh in in vitro bacterial cultures analysed in d. f, CHRM4 activation by diverse Bifidobacterium strains cultured in vitro with 1 mM choline. In b–f, CHRM4 activity data from in vitro samples are representative of three independent experiments (n = 3 biologically independent samples per group in each experiment); CHRM4 activity data from in vivo samples are representative of two independent experiments (n = 3 mice per group in each experiment). ACh quantification data are from one representative independent experiment. Data are mean + s.e.m in a, and mean ± s.e.m in b–f. Statistical significance was assessed using one-way ANOVA followed by Tukey’s post hoc test (b (in vitro)) and Welch’s ANOVA followed by Games–Howell post hoc test for (b (in vivo), d and f). In b–f, statistical comparisons were performed between each bacterial in vitro culture and the medium control, and between monocolonized groups and the GF control. Exact P values are shown in the figure when P < 0.05; all other P values are provided in the Source data. n.d., not detected.Source dataWe selected a representative strain from each phylum to validate the observed CHRM4 activity using CHRM4-Tango: B. longum NWP308 (Actinobacteria), B. caccae NWP468 (Bacteroidetes), P. pentosaceus NP38 (Firmicutes) and Bacteroides ovatus NWP195 as a non-CHRM4-activating control. We monocolonized groups of GF mice with each strain and prepared caecal metabolome libraries 2 weeks after colonization. Caecal metabolomes from mice colonized with putative CHRM4-activating strains triggered CHRM4 activation, while metabolomes from GF or B. ovatus NWP195-colonized mice lacked this activity (Fig. 2b). Consistent with our initial screening results, in vitro metabolomes from all strains failed to activate CHRM4 (Fig. 2b). Mass spectrometry analysis of caecal contents from P. pentosaceus NP38, B. longum NWP308 and B. caccae NWP468-colonized mice revealed high levels of ACh, whereas caecal contents from B. ovatus NWP195-colonized and GF mice showed no detectable ACh (Fig. 2c).Commensal strains that produce ACh in vivo may lack this activity in vitro due to an absence of required chemical precursors in standard culture medium. Notably, all of the in vivo CHRM4-active bacteria identified in our original screen were cultured in gut microbiota medium (GMM)14, which lacks the essential dietary nutrient and ACh precursor choline (Supplementary Table 4). Accordingly, supplementation of GMM with 1 mM choline restored robust ACh production and CHRM4 activation in all tested strains aside from the negative control strain B. ovatus NWP195, with P. pentosaceus NP38 showing the highest activity (Fig. 2d,e). These results indicate that the ACh biosynthetic pathway is functional in vitro but limited by substrate availability under standard culture conditions. Consistent with this, the standard mouse diet used in our facility contains approximately 1,200 mg per kg choline, providing an abundant source of choline to commensal microorganisms grown in vivo.We noted that multiple Bifidobacterium species, which dominate the infant gut microbiota15,16,17 (Supplementary Fig. 2a,b), activated CHRM4 in our screen. To broadly explore ACh synthesis across different bifidobacteria, we tested 13 representative Bifidobacterium strains from our human gut commensal collections for CHRM4 activation. When cultured in the presence of choline, all B. breve strains, B. longum strains and one Bifidobacterium dentium strain activated CHRM4, with B. breve NWP289 showing the strongest activity (Fig. 2f). B. breve and B. longum are among the most prevalent Bifidobacterium strains in infant microbiomes, while B. dentium is less prevalent but is highly abundant in some individuals15 (Supplementary Fig. 2c). Human breast milk is enriched in choline-containing compounds (approximately 1,400 μM total choline, including 140 μM free choline)18. All tested Bifidobacterium strains converted free choline into ACh; by contrast, most Bifidobacterium strains did not efficiently convert glycerophosphocholine, phosphocholine or sphingomyelins into ACh, although B. dentium NWP234 showed evidence of phosphatidylcholine utilization (Supplementary Fig. 3). Together, these data suggest that infant microbiomes are dominated by ACh-producing Bifidobacterium species exposed to high levels of choline in breast milk or formula, creating a favourable environment for high-level production of microbiota-derived ACh. This prompted us to examine the mechanisms by which commensal species produce ACh and the potential impacts of microbial ACh production on host biology.Mechanisms of commensal ACh synthesisACh synthesis in mammals is catalysed by choline acetyltransferase (ChAT), which transfers an acetyl group from acetyl-CoA to the hydroxyl substituent of choline (Extended Data Fig. 6a). We could not identify any homologues of mammalian ChAT in the B. breve NWP289 genome. Thus, we next searched for acetyltransferases using rapid annotations using subsystems technology (RAST)19 and identified 14 full-length putative acetyltransferases (Supplementary Table 5). Nine sequences annotated as N-acetyltransferases were excluded from further study. We cloned and recombinantly expressed the other five putative acetyltransferases. After purification (Extended Data Fig. 6b), we tested their ability to catalyse ACh synthesis from acetyl-CoA and choline in vitro (Extended Data Fig. 6c). Products from putative acetyltransferase 2 exhibited potent CHRM4 activity and high ACh levels, while the products of putative acetyltransferases 1 and 5 exhibited only very weak activity and no detectable ACh (Fig. 3a,b). Thus, putative acetyltransferase 2 is the primary enzyme that catalyses ACh synthesis in B. breve NWP289; hereafter, we refer to this enzyme as B. breve ChAT (BbChAT).Fig. 3: Gut microorganisms use hexapeptide repeat proteins to synthesize ACh.The alternative text for this image may have been generated using AI.Full size imagea,c, CHRM4 activation by products from in vitro enzyme assays using recombinant putative acetyltransferases from B. breve NWP289 (a) or P. pentosaceus NP38 (c) incubated with choline and acetyl-CoA. b,d, Mass spectrometry quantification of ACh in the assay products from a and c, respectively. e, AlphaFold2 models of BbChAT and PpChAT, and crystal structure of EcMoAT (PDB: 1OCX). f, Sequence alignment of hexapeptide repeat acetyltransferase family proteins showing the conservation of the catalytic histidine. B. anthracis, Bacillus anthracis; C. difficile, Clostridioides difficile; G. kaustophilus, Geobacillus kaustophilus; S. aureus, Staphylococcus aureus. g, CHRM4 activation by products from WT and mutant BbChAT and PpChAT assayed under the same conditions as in a and c. h, Phylogenetic tree of the prokaryotic hexapeptide repeat protein superfamily. i, CHRM4 activation by assay products from selected hexapeptide repeat proteins (highlighted in h). j, CHRM4 activation by WT and Δchat B. breve cultured with and without 1 mM choline. k, Mass spectrometry quantification of ACh in cultures of WT and Δchat B. breve grown with 1 mM choline. CHRM4 activity (a, c, g, i and j) and ACh quantification (b, d and k) are representative of two independent experiments (n = 3 biologically independent samples per group in each experiment). Data are mean ± s.e.m. Statistical significance was assessed using Welch’s ANOVA with Games–Howell post hoc test (a, c, d and i), Kruskal–Wallis tests followed by Dunn’s multiple-comparison test (b), two-sided Welch’s t-tests (g) and one-way ANOVA with Tukey’s post hoc test (j). Statistical comparisons were performed between each candidate enzyme and the no-protein control (a–d and i), and between each bacteria culture and the medium control (j). Exact P values are shown in the figure when P < 0.05; all other P values are provided as Source data.Source dataWe similarly analysed the P. pentosaceus NP38 genome and selected five candidate acetyltransferases for characterization (Supplementary Table 5). We cloned and purified all selected enzymes and evaluated their ability to produce ACh and activate CHRM4 in vitro (Extended Data Fig. 6d). Among these enzymes, putative acetyltransferase 4 (hereafter PpChAT) demonstrated the highest activity, while putative acetyltransferase 5 showed only very mild activity (Fig. 3c,d). To better characterize these two ACh-producing enzymes, we modelled their structures using AlphaFold2 and searched for homologues within the Protein Data Bank (PDB; Fig. 3e). While BbChAT and PpChAT share only approximately 30% amino acid identity, structural predictions revealed that they both belong to the hexapeptide repeat acetyltransferase family, showing high structural homology to Escherichia coli maltose O-acetyltransferase (EcMoAT; DALI Z score: PpChAT, 29.6; BbChAT, 26.4; Fig. 3e), another member of the hexapeptide repeat acetyltransferase family that has been structurally resolved20,21. Alignment of BbChAT and PpChAT revealed conservation at the putative active site histidine (Fig. 3f), which was required for ACh synthesis by these enzymes (Fig. 3g and Extended Data Fig. 6b,e).The hexapeptide repeat protein superfamily contains hundreds of thousands of sequences in InterPro. To contextualize the commensal-encoded ChAT enzymes identified here, we generated a phylogenetic tree of all prokaryotic hexapeptide repeat proteins in the PDB with experimentally determined structures (Fig. 3h). These proteins were primarily annotated as acetyltransferases but varied widely in their predicted substrate specificity21. Proteins clustered in the same clade generally had related annotated functions, for example, cell wall biosynthesis, antibiotic resistance, L-lysine biosynthesis, L-cysteine biosynthesis and lipid-A biosynthesis (Fig. 3h). PpChAT and BbChAT localized to a clade of maltose/galactoside acetyltransferases that also included EcMoAT and E. coli LacA.To examine the potential ChAT activities of proteins in this superfamily, we selected three additional proteins from the maltose/galactoside acetyltransferase clade and one protein each from the neighbouring antibiotic resistance and L-lysine biosynthesis clades (Supplementary Table 5) and assessed their ability to convert choline to ACh as measured on the basis of CHRM4 activation. The newly described ChATs from Bifidobacterium or Pediococcus exhibited the strongest activity, while the LacA (E. coli) and CatB8 (Acinetobacter baumannii) samples exhibited very weak CHRM4 activation, and MoAT (E. coli), DAPD (Enterococcus faecalis) and HrcAT (Vibrio cholerae) exhibited minimal activity (Fig. 3i), showing that ChAT activity is unique and not a universal feature of these related maltose/galactoside acetyltransferases or hexapeptide repeat proteins more broadly.Given that BbChAT was the most active ChAT from B. breve NWP289, we next examined whether this gene was required for B. breve ACh biosynthesis. We constructed a BbChAT mutant (B. breve::pFREM28-Bb_chat, hereafter Δchat B. breve), in which the coding sequence of BbChAT was disrupted (Extended Data Fig. 6f–h). We then cultured wild-type (WT) B. breve NWP289 (hereafter WT B. breve) and Δchat B. breve in the presence or absence of 1 mM choline. Δchat B. breve did not produce ACh or activate CHRM4 (Fig. 3j,k), demonstrating that BbChAT is essential for ACh synthesis in B. breve NWP289.Commensal ACh enhances intestinal IgATo test the potential impacts of commensal ACh on gut physiology, we monocolonized GF mice with WT or Δchat B. breve and then treated these animals with or without the FDA-approved acetylcholinesterase inhibitor (AChEi) rivastigmine to enhance the potential effects of commensal ACh. WT and Δchat B. breve strains colonized the gut at similar levels both in the presence and absence of AChEi under monocolonization conditions (Extended Data Fig. 7a,b); furthermore, consistent with our previous bioactivity-based assessments, luminal contents from WT B. breve-colonized mice contained approximately 6 µg per g ACh, whereas ACh was undetectable in Δchat B. breve-colonized mice (Extended Data Fig. 7c,d). As the major parasympathetic neurotransmitter, ACh regulates gut motility and epithelial fluid secretion22. Accordingly, we observed that mice colonized with WT B. breve and treated with AChEi exhibited significantly increased gut motility compared with those colonized with the Δchat strain (Fig. 4a), whereas this difference was blunted without AChEi supplementation (Extended Data Fig. 7e). B. breve-induced increases in gut motility were abolished by treatment with the CHRM3 antagonist 4-DAMP (Fig. 4a), indicating that bacterial-derived ACh can increase gut motility through a CHRM3-dependent mechanism. By contrast, we did not observe any measurable differences in faecal water content between WT and Δchat B. breve-colonized mice (Extended Data Fig. 7f,g).Fig. 4: ACh-producing B. breve increases gut motility and enhances intestinal IgA responses.The alternative text for this image may have been generated using AI.Full size imagea, Whole-gut transit time of WT or Δchat B. breve-monocolonized mice treated with AChEi, with or without 4-DAMP (1 mg per kg, intraperitoneal (i.p.)). n = 5, 4, 5 and 4 mice per group. b, Schematic of bulk RNA-seq of LP immune cells from WT or Δchat B. breve-monocolonized mice treated with AChEi. c, Differentially expressed genes in LP immune cells (DESeq2, two-sided Wald tests with Benjamini–Hochberg correction). d, GSEA of differentially expressed genes. e–h, Representative flow cytometry plots (e) and the frequency (f) and number (g) of colonic LP IgA plasma cells, and faecal IgA concentrations (h) in monocolonized mice. n = 8 and 7 (f and g) and n = 8 (h) mice per group. i, Schematic of colonization of antibiotic-treated SPF mice with WT or Δchat B. breve. j–m, Representative flow cytometry plots (j) and the frequency (k) and number (l) of colonic LP IgA plasma cells, and faecal IgA concentrations (m) in colonized SPF mice. n = 5 mice per group. n–q, Representative flow cytometry plots (n and p) and the frequency of IgA-coated bacteria (o and q) in monocolonized mice (n and o; n = 8 mice per group) and colonized SPF mice (p and q; n = 5 mice per group). r–u, Representative flow cytometry plots (r) and the frequency (s) and number (t) of colonic LP IgA plasma cells, and faecal IgA concentrations (u) in Δchat and WT B. breve-colonized mice treated with atropine or mecamylamine (25 mg l−1 each in drinking water). n = 5, 4, 4 and 4 mice per group. Data are mean ± s.e.m. and representative of two (a, j–m and p–u) or three (e–h, n and o) independent experiments. Statistical significance was assessed using unpaired two-sided t-tests (a, f–h, k–m, o and q) and one-way ANOVA followed by Tukey’s post hoc test (s–u). The diagrams in b and i were created using BioRender; Song. D https://BioRender.com/8t518es (b) and https://BioRender.com/vhqblos (i) (2026).Source dataACh has also been shown to modulate innate and adaptive immunity23,24,25,26,27,28. To examine the impacts of commensal ACh on the mucosal immune system, we performed single-cell RNA sequencing (scRNA-seq) analysis of gut lamina propria (LP) immune cells from mice monocolonized with WT and Δchat B. breve. We did not observe any marked differences in cellular frequencies; however, ileal plasma cell and naive/memory B cell populations exhibited transcriptional differences in mice colonized with WT versus Δchat B. breve (Extended Data Fig. 8a). Pseudobulk DESeq2 analysis revealed increased Igha gene expression in plasma cells and naive/memory B cells from mice colonized with WT versus Δchat B. breve (Extended Data Fig. 8b–g); increased Igha expression was also apparent in a reclustered UMAP of plasma cell, naive/memory B cell and germinal centre B cell subsets (Extended Data Fig. 8h). Next, we treated monocolonized mice with AChEi to enhance the potential effects of commensal ACh and performed bulk RNA-seq analysis of colonic and ileal LP immune cells (Fig. 4b). Mice colonized with WT B. breve showed significantly higher colonic expression of Aicda, Igha, Jchain, Mzb1, Bhlha15, Tnfrsf17, Mef2b and Ccl21d, compared with those colonized with Δchat B. breve (Fig. 4c). Ileal transcripts exhibited similar trends (Fig. 4c and Supplementary Fig. 4). Gene set enrichment analysis (GSEA) revealed that colonic LP immune cells from mice colonized with WT B. breve were enriched for pathways related to immunoglobulin production, immunoglobulin-mediated immune response, B-cell-mediated immunity, production of molecular mediator of immune response and adaptive immune response (Fig. 4d).We next analysed colonic LP immune cells using flow cytometry (Supplementary Fig. 5a). Consistent with the transcriptomic data, WT B. breve colonization led to a higher frequency and absolute number of plasma cells in the colonic LP (Fig. 4e–g). Similarly, we observed a significant increase in faecal IgA levels in WT versus Δchat B. breve-colonized mice (Fig. 4h). To assess whether this effect was evident in a complex microbial context, we colonized specific-pathogen free (SPF) mice with WT or Δchat B. breve by pretreating SPF mice with antibiotics and then gavaging with B. breve for 5 weeks to establish stable colonization (Fig. 4i). Consistent with our monocolonization results, colonization with WT B. breve induced increased colonic plasma cell frequency and faecal IgA levels as compared to colonization with Δchat B. breve (Fig. 4j–m). However, in this context, the Δchat strain exhibited significant colonization defects compared with the WT control (Δchat, around 1% relative abundance; WT, around 11% relative abundance) (Extended Data Fig. 7h). These results suggest that ACh-producing B. breve can promote enhanced IgA production and that B. breve ChAT increases bacterial fitness in the presence of a complex SPF microbial community.IgA is secreted into the gut lumen, where it binds to commensal bacteria at steady state29,30,31. WT mice monocolonized with B. breve and treated with or without AChEi exhibited a higher proportion of IgA-coated faecal bacteria than Δchat-colonized mice (Fig. 4n,o, Extended Data Fig. 9a,b and Supplementary Fig. 5b). We observed similar effects in SPF mice colonized with WT or Δchat B. breve (Fig. 4p,q). To assess the specificity of these IgA responses, we stained in vitro-grown B. breve NWP289 and ten unrelated bacterial species with faecal IgA from mice monoassociated with WT or Δchat B. breve (Extended Data Fig. 9c). We observed limited faecal IgA binding to in vitro-grown non-B. breve strains and, notably, also to in vitro-grown B. breve across all groups (Extended Data Fig. 9d–f). On the basis of these data, we hypothesized that in vitro- and in vivo-grown B. breve may differ in the cell-surface structures targeted by IgA. To test this possibility, we monocolonized Rag1-deficient (Rag1−/−) mice with B. breve and used faecal bacteria from these mice as a source of non-antibody-bound in vivo-grown B. breve. We found that concentration-normalized faecal IgA from WT B. breve-colonized mice bound to in vivo-cultivated B. breve at a higher level than IgA from Δchat B. breve-colonized mice, while binding to a defined community of non-B. breve strains grown in Rag1−/− mice was similar between groups (Extended Data Fig. 9g–l). Together, these data suggest that ACh-producing B. breve preferentially enhances the production of B. breve-reactive IgA.Finally, to discern whether muscarinic versus nicotinic ACh receptors mediated B. breve-induced IgA enhancement, we supplemented atropine (a pan-muscarinic ACh receptor antagonist) or mecamylamine (a pan-nicotinic ACh receptor antagonist) in the drinking water of mice monoassociated with B. breve. Mecamylamine treatment abolished WT B. breve-induced increases in colonic plasma cells and faecal IgA levels, whereas atropine had no significant effect (Fig. 4r–u). These findings suggest that ACh-producing B. breve enhances intestinal IgA responses primarily through nicotinic ACh receptors.ACh+
Commensal-derived acetylcholine enhances mucosal immune education - Nature
A diet–microbiome–host axis strengthens mucosal immune defences and reinforces host–microbiota mutualism.
15,629 words~71 min read