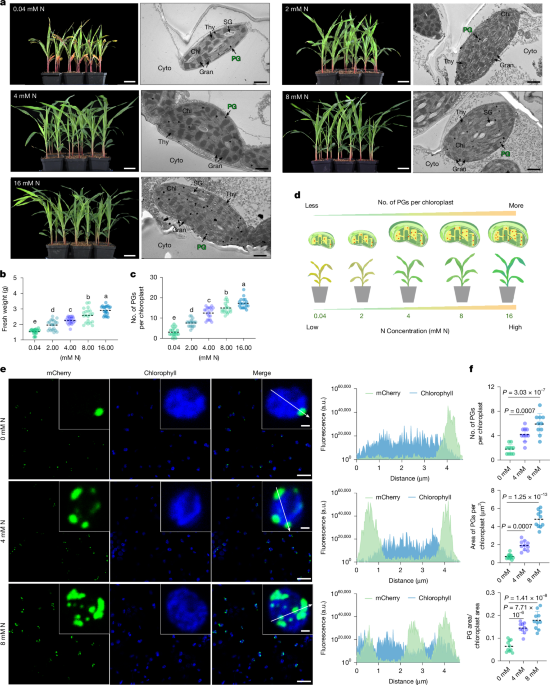
MainNitrogen (N) is essential for plant growth and agricultural productivity, directly underpinning global food security. However, improving crop nitrogen use efficiency (NUE) remains a global challenge. Approximately 70% of applied N fertilizer is inefficiently used, with maize NUE often below 30% (ref. 1). This inefficiency causes economic burdens and environmental degradation. Enhancing crop NUE is an urgent imperative. Maize is a globally important cereal crop for food, feed and industry2. As a highly domesticated C4 plant, maize exhibits a strong physiological interdependence among its growth, development and nitrogen utilization3. Many agricultural systems worldwide are at present grappling with the dual challenges of yield dependence on increased nitrogen input and inherently low NUE, highlighting an urgent need for fundamental breakthroughs in nitrogen utilization mechanisms in crops4,5.Maize NUE is a complex trait influenced by nitrogen absorption, assimilation and remobilization. Nitrogen assimilation is important for converting inorganic nitrogen into organic forms, directly affecting growth and yield. Although plants primarily absorb nitrate (NO3−) and ammonium (NH4+), the subcellular organization of this pathway remains a key determinant of efficiency. Maize, a typical C4 plant, possesses Kranz anatomy in its leaves, characterized by concentric layers of outer mesophyll cells (MCs) and inner bundle sheath cells (BSCs) that tightly encase the vascular bundles. Previous studies have shown that the key enzymes for primary nitrogen assimilation, nitrate reductase (NR) and nitrite reductase (NIR), are localized exclusively in MCs6. By contrast, although glutamine synthetase (GLN, also known as GS) is present in both cell types, ferredoxin-dependent glutamate synthase (Fd-GOGAT) is almost entirely confined to the chloroplasts of BSCs7. Consequently, the proposed model for the cellular compartmentalization of nitrogen assimilation suggests that, during primary nitrogen assimilation, MCs harbouring NR, NIR and GLN play an important part in synthesizing glutamine from NO3−. Following root uptake and transport to the leaves by the vasculature, NO3− diffuses into MCs. In the cytoplasm, NR reduces NO3− to nitrite (NO2−) using nicotinamide adenine dinucleotide (NADH) as an electron donor8,9. Subsequently, NO2− is transported into chloroplasts, where NIR reduces it to NH4+ using reduced Fd supplied by the photosynthetic electron transport chain. GLN then converts the free NH4+ into glutamine (Gln), thereby facilitating the conversion of inorganic to organic nitrogen10,11. Concurrently, the high level of Fd-GOGAT present in BSCs may drive substantial glutamate synthesis, completing the GS-GOGAT cycle required for the synthesis of amino acids and other organic compounds12. It is important to note that NO2− and NH4+ are intermediates in early nitrogen assimilation; their accumulation to toxic levels can damage plant cells13. Therefore, the spatial organization provided by cellular compartmentalization is crucial for ensuring orderly nitrogen assimilation, facilitating both rapid nitrogen processing and the detoxification of metabolic intermediates. Although mutant analyses and quantitative trait locus cloning have identified several nitrogen utilization genes, such as nitrate transporter1.1B (NRT1.1B), GROWTH-REGULATING FACTOR 4 (GRF4), TEOSINTE-BRANCHED1/CYCLOIDEA/PCF 19 (TCP19), NITROGEN-MEDIATED TILLER GROWTH RESPONSE 5 (NGR5) in rice (Oryza sativa L.) and TEOSINTE HIGH PROTEIN 9 (THP9) in maize, and validated their roles in enhancing NUE14,15,16,17,18, the patterns of nitrogen assimilation within internal subcellular compartments, as well as the core mechanisms regulating them, need to be explained.Cells contain various distinct, compartmentalized structures, including organelles and membrane-less biomolecular condensates, which are essential for survival and the efficient execution of biological functions. These compartments provide spatial organization and serve as independent microenvironments for biochemical reactions, thereby facilitating the orderly and efficient progression of intracellular physiological and metabolic processes19. Plastoglobules (PGs) are lipid droplet-like structures located on the thylakoid membranes, enclosed by a lipid monolayer20,21,22. Current research indicates that PGs serve not only as central sites for lipid storage and metabolism but also as dynamic subcompartments that respond to diverse environmental stimuli22,23. Factors such as drought, temperature, salinity, light and nutrient deficiency can influence the abundance and size of PGs24,25,26,27,28. Here, we show that PGs exhibit striking nitrogen-responsive dynamics. This phenomenon offers new insights into nitrogen metabolic mechanisms in maize, redefining the role of PGs in plant primary metabolism and providing a new framework for enhancing crop productivity through deep mechanistic and structural insights.PGs exhibit nitrogen-responsive dynamicsTo investigate the nitrogen response mechanisms of maize, we conducted a nitrogen gradient experiment using the B73 inbred line. We evaluated phenotypic traits and physiological parameters of maize seedlings under five distinct nitrogen concentrations (0.04 mM, 2 mM, 4 mM, 8 mM and 16 mM N/KNO3). The results showed a strong positive correlation between biomass accumulation and nitrogen availability (Fig. 1a,b). Plants subjected to the low nitrogen (LN, 0.04 mM N) condition exhibited a characteristic yellowish hue, which progressively deepened to green as nitrogen availability increased (Fig. 1a). These findings are consistent with nitrogen response phenotypes previously reported in other crops29.Fig. 1: Nitrogen availability influences both the number and size of PGs.The alternative text for this image may have been generated using AI.Full size imagea, Phenotypes of maize seedlings grown under five distinct nitrogen concentrations (0.04 mM, 2 mM, 4 mM, 8 mM and 16 mM N/KNO3) and electron microscope images of chloroplasts in their MCs. Scale bar, 5 cm (phenotype images); 1 µm (electron microscope images). Chl, chloroplast; Cyto, cytoplasm; SG, starch grain; Gran, granum; Thy, thylakoid; and PG, plastoglobule. b, Quantification of fresh weight of maize seedlings under five distinct nitrogen concentrations. Data are mean ± s.d. (n = 20 independent biological plants harvested from a single, randomized experiment). c, Quantification of PG number per chloroplast in MCs under five distinct nitrogen treatments. Data are mean ± s.d. (n = 18 independent biological chloroplasts from a single, randomized experiment). d, Schematic of maize nitrogen response to varying nitrogen concentrations, highlighting the dynamic changes in PGs. e,f, Representative fluorescence images (e), quantification of PG number (f, top), PG area per chloroplast (f, middle) and the ratio of PG area to chloroplast area (f, bottom). Maize PG-marker protein ZmPSY3 fused to the mCherry fluorescent tag was transiently expressed in tobacco leaf epidermal cells, followed by treatment with nitrogen solutions at different concentrations. Observations were performed on at least 10 cells from three independently transformed tobacco leaves, and the size and number of PGs were quantified by fluorescence microscopy. Scale bar, 1 µm (partial images), 20 µm (complete images). In f, the data are mean ± s.d. (n = 10 biological replicates for counting PG number per chloroplast, n = 10 biological replicates for measuring PG area per chloroplast), In b,c, different letters denote significant differences (P < 0.05) based on one-way ANOVA with Tukey’s test; see Source Data. In f, two-tailed Student’s t-tests were used to determine P values; see Source Data. a.u., arbitrary units.Source dataGiven that leaves are primary sites for photosynthesis and nitrogen assimilation30, we performed transmission electron microscopy (TEM) on maize leaves under varying nitrogen conditions. We imaged and analysed the chloroplasts of MCs and BSCs separately (Fig. 1a and Extended Data Fig. 1c). Notably, we observed a progressive increase in the number of PGs in MC chloroplasts that correlated with rising nitrogen levels in MC chloroplasts (Fig. 1c). This dynamic change in PGs was not attributable to an increase in MC chloroplast size (Extended Data Fig. 1a,b). By contrast, this phenomenon was not observed in BSC chloroplasts, in which PG numbers showed no nitrogen-dependent trend, and the chloroplast size remained consistent across different nitrogen concentrations (Extended Data Fig. 1d,e). The relatively low abundance of PGs in BSC chloroplasts under 0.04 mM and 2 mM nitrogen treatments is probably attributed to reduced thylakoid formation under LN conditions, which limits the membrane area available for PG budding, rather than reflecting a direct decrease in PG biogenesis. Collectively, these results indicate that nitrogen not only influences biomass accumulation and leaf colouration at the whole plant level but also modulates PG dynamics in MC chloroplasts at the subcellular level (Fig. 1d).To further explain the dynamic behaviour of PGs in response to nitrogen, we transiently expressed a PG-marker protein fused to mCherry31, ZmPSY3 (phytoene synthase 3)-mCherry, in tobacco leaves. Fluorescence microscopy showed pronounced nitrogen-dependent alterations in both the morphology and quantity of PGs. Compared with N-free treatment (0 mM N), both the number and size of PGs markedly increased at 4 mM nitrogen treatment. At 8 mM nitrogen, PG number and size further increased, accompanied by the formation of morphologically irregular aggregates (Fig. 1e,f). These results unequivocally demonstrate that chloroplast-localized PGs exhibit highly dynamic behaviours in response to nitrogen in maize.PG dynamics in C3 and C4 plantsTo examine whether the dynamic responses of PGs to nitrogen are conserved in plants, we extended our investigation to C3 plant species, including the dicots soybean (Glycine max (L.) Merr.) and tobacco (Nicotiana tabacum L.), as well as two monocots, rice and wheat (Triticum aestivum L.). Each species was subjected to two nitrogen treatments. Consistent with our observations in maize MC chloroplasts, all four tested species exhibited notable increases in both biomass and chlorophyll content under higher nitrogen levels (Extended Data Fig. 1f–h). Notably, PGs in these species exhibited dynamic responses to nitrogen availability remarkably similar to those observed in maize, with PG numbers increasing concomitantly with nitrogen levels (Extended Data Fig. 1i,j). These findings establish nitrogen-induced PG dynamics in MC chloroplasts as a conserved and fundamental response across both C3 and C4 plant lineages.Maize PG fractionation and profilingTo delve into the biochemical basis of nitrogen responsiveness of maize PGs, we successfully adapted and optimized the Arabidopsis thaliana PG isolation methodology (Extended Data Fig. 2). To separately isolate PGs from MC and BSC chloroplasts, we used differential mechanical disruption using a blender at varying power settings32. MCs are effectively detached under low-power blending, whereas BSCs require multiple rounds of high-power blending. We processed 200 g of fresh leaf material, yielding four batches of highly concentrated MC chloroplasts and one batch of BSC chloroplasts (Extended Data Fig. 2a). Immunoblot analysis using cell-type-specific markers confirmed the purity of the separation, in which PEPC was detected exclusively in MCs, whereas RuBisCO was restricted to BSCs (Extended Data Fig. 2b).Our refined protocol involved initial isolation of intact MC chloroplasts, followed by ultrasonic disruption to release PGs and membrane fragments. The resulting homogenates were then subjected to two rounds of sucrose gradient ultracentrifugation to yield highly purified PGs. To assess the purity of the extracted PGs, we used specific organelle markers: tocopherol cyclase (VTE1) for PGs, TOC75 for chloroplast outer envelope membrane and chlorophyll a/b-binding LHCB1 for the thylakoid membrane33,34. These analyses confirmed successful isolation of relatively pure PGs from MC chloroplasts (Extended Data Fig. 2c). The purified PGs were then subjected to trypsin digestion and desalting, and the resulting peptides were analysed by liquid chromatography–tandem mass spectrometry (LC-MS/MS). Proteomics data were acquired by data-independent acquisition (DIA) technology (Fig. 2a). This rigorous approach allowed us to obtain comprehensive PG proteomic profiles from two biological replicates of B73 inbred leaves.Fig. 2: Purification and validation of maize PG components.The alternative text for this image may have been generated using AI.Full size imagea, Schematic of maize PG protein extraction and purification. b, Heatmap showing the abundance of representative proteins in PGs (left) and the log 2 of protein intensities in PGs (right). The bar graph shows the mean of two independent biological replicates (B73-1 and B73-2). S-B metabolism, secondary-biosynthetic metabolism; C–N metabolism, carbon–nitrogen metabolism. c, Subcellular localization of representative proteins shown in b. Representative fluorescence images of eGFP-tagged proteins in tobacco epidermal cells are shown. PSY3-mCherry, a PG-localized marker. Scale bar, 1 µm. Partial elements of a (Export ID: OAWTTe6e6c) were drawn using the FigDraw 2.0 platform (https://www.figdraw.com/).Source dataNitrogen assimilation enzymes in PGsWe conducted a functional proteomic classification of the highly abundant proteins in maize PGs. These proteins were primarily categorized into three groups: structural proteins, enzymes involved in secondary metabolic pathways and proteins involved in carbon–nitrogen metabolic processes. Consistent with previous findings in maize and other plant species35,36,37,38,39,40,41, our proteomic data identified five fibrillin proteins, including putative plastid-lipid-associated protein 1 chloroplastic (ZmPLAP1), ZmPLAP10, ZmPLAP13, ZmPLAP2 and plastid-lipid-associated protein 2 (ZmB4FRP8). These fibrillins, known to exhibit high abundance, constitute a predominant protein component within PGs. The second category of PG proteins includes enzymes involved in secondary metabolite biosynthesis, such as sucrose export defective 1 (ZmSDX1), nine-cis-epoxycarotenoid dioxygenase 6 (ZmNCED6), lipoxygenase 10 (ZmLOX10) and lemon white 2 (ZmLW2). This is consistent with previous studies demonstrating the role of PGs lipid metabolism, carotenoid biosynthesis and phenolic compound synthesis23,35.Our analysis identified two key nitrogen assimilation enzymes, ZmNIR2 and ZmGLN1, as integral components of maize PGs (Fig. 2b). We also detected carbon-metabolism-associated proteins of particular significance, namely, carbonic anhydrase 1 (ZmCAH1) and carbonic anhydrase 6 (ZmCAH6). Subcellular localization analyses of representative proteins from each category confirmed their co-localization with an mCherry-fused PG marker protein in tobacco leaves (Fig. 2c and Supplementary Figs. 1 and 2), providing strong evidence for their presence within PGs.PG localization and vegetative growthThe identification of ZmNIR2 and ZmGLN1 as integral components of maize PGs (Fig. 2b) prompted us to investigate the functional significance of their specific subcellular localization. Transcriptomic data from qTeller MaizeGDB showed that among the two ZmNIR genes (ZmNIR1 and ZmNIR2) and six ZmGLN genes (ZmGLN1–6) in the maize genome, ZmNIR2 and ZmGLN1 are predominantly expressed in leaves (Fig. 3a), a primary site for nitrogen assimilation. This expression pattern aligns with their potential role in leaf-centric nitrogen metabolism. To further explore the functional relationship between PG localization and the roles of ZmNIRs and ZmGLNs, we generated eight single mutant lines: nir1, nir2-1, nir2-2, gln1, gln2, gln3, gln4, gln5 and gln6 mutants (Extended Data Fig. 3). Phenotypic analyses showed that ZmNIR2 mutants exhibited severe stunted growth and leaf yellowing, whereas gln1 exhibited reduced plant height and prolonged vegetative growth (Fig. 3b,d and Extended Data Fig. 3). Notably, ZmNIR2 mutants showed chlorotic phenotype even under sufficient nitrogen supply. By contrast, nir1 and gln3-6 mutants showed a normal phenotype. Although gln2 exhibited reduced plant height, there was no significant difference in biomass (Fig. 3c,d).Fig. 3: PG localization is important for the function of nitrogen assimilation enzymes.The alternative text for this image may have been generated using AI.Full size imagea, Transcript levels of maize ZmNIR genes (ZmNIR1 and ZmNIR2) and ZmGLN genes (ZmGLN1−6) in root, stem, leaf and reproductive tissues. b,c, Representative phenotypes of nir1, nir2-1, gln1, gln2, gln3, gln4, gln5 and gln6 mutants and their corresponding controls (WT/CK). Scale bar, 20 cm. d, Quantification of fresh weight and plant height for nir1, nir2-1, gln1, gln2, gln3, gln4, gln5 and gln6 mutants and their corresponding controls. Data are presented as mean ± s.d. (n = 3 independent biological plants for fresh weight and n = 10 independent biological plants for plant height; all data were harvested from a single, randomized experiment). Two-tailed Student’s t-tests were used to determine the P values. Different letters denote significant differences (P < 0.05) based on one-way ANOVA with Tukey’s test. e, Representative fluorescence images of ZmNIR1-eGFP, ZmNIR2-eGFP, ZmGLN1-eGFP, ZmGLN2-eGFP, ZmGLN3-eGFP, ZmGLN4-eGFP, ZmGLN5-eGF, and ZmGLN6-eGFP proteins in tobacco epidermal cells, co-localized with the PG-localized marker PSY3-mCherry. Scale bar, 1 µm (partial images), 20 µm (complete images).Source dataTo precisely determine the subcellular localization of the proteins encoded by these eight genes, we performed detailed localization assays. Our findings unequivocally demonstrated that only ZmNIR2 and ZmGLN1 are uniquely targeted to PGs, co-localizing with the mCherry-fused PG marker protein in tobacco leaves. By contrast, ZmNIR1 was predominantly localized to the chloroplast, and ZmGLN2-6 were consistently localized to the cytoplasm (Fig. 3e), which is consistent with previous observation13,42. Notably, we also observed a minor fraction of ZmNIR1 protein localized to PGs, indicating weaker PG-targeting ability (Extended Data Fig. 4a). However, marked differences remain between ZmNIR1 and ZmNIR2. ZmNIR1 is primarily expressed in roots, whereas ZmNIR2 is highly expressed in leaves (Extended Data Fig. 4b). Regarding relative protein abundance within PGs, ZmNIR1 was detected at only around 2,000 compared with approximately 200,000 for ZmNIR2 (Extended Data Fig. 4c). This disparity probably explains why loss of ZmNIR1 has a far less pronounced impact than that of ZmNIR2.PG localization is key for maize NUETo further investigate the roles of ZmNIRs and ZmGLNs in nitrogen response, we conducted a nitrogen gradient experiment using the eight mutant lines under three distinct nitrogen treatments (KCl, 0 mM N; LN, 0.04 mM N; NN, 4 mM N/KNO3). Phenotypic analyses showed that the nir2-1 mutant showed no biomass difference from WT under KCl, suggesting a specific response to N availability. Under LN, WT fresh weight increased, whereas nir2-1 showed no change. Under NN, nir2-1 fresh weight was reduced to 57% of WT with leaf yellowing (Extended Data Fig. 5a). The gln1 mutant consistently showed reduced fresh weight across all N concentrations, reaching 51% of WT under KCl and 60% under NN (Extended Data Fig. 5c). Conversely, nir1 and gln2-6 mutants showed no defects (Extended Data Fig. 5b,d). These experiments confirm the important, non-redundant roles of ZmNIR2 and ZmGLN1 in nitrogen response. These experiments together confirm the important and non-redundant roles of ZmNIR2 and ZmGLN1 in nitrogen response. Mutants lacking these specific, PG-localized enzymes exhibit strongly reduced sensitivity to nitrogen and impaired NUE.To investigate whether the subcellular localization of ZmNIRs and ZmGLNs exhibits dynamic changes in response to nitrogen, we treated tobacco cells transiently expressing ZmNIRs-eGFP and ZmGLNs-eGFP by infiltration with solutions of three nitrogen concentrations (0 mM, 4 mM and 8 mM). The results show that ZmNIR2 and ZmGLN1 exhibit an increase in PG-localization with rising nitrogen concentrations, whereas the other ZmGLNs and ZmNIR1 show no change in their subcellular localization patterns (Extended Data Fig. 6).Hydrophobic-region-mediated PG targetingThe chloroplast targeting mechanisms and the PG-targeting mechanisms of ZmNIR2 and ZmGLN1 are two independent processes. As chloroplast-targeted proteins ZmNIR2 and ZmGLN1 typically contain an N-terminus cTP. Using DeepLoc-2.1 (https://services.healthtech.dtu.dk/services/DeepLoc-2.1/), the predicted cTP for ZmNIR2 spans amino acids 1–47, and for ZmGLN1, amino acids 1–51. TargetP-2.0 (https://services.healthtech.dtu.dk/services/TargetP-2.0) predicts the cleavage site of the ZmNIR2 cTP to be between amino acids 37 and 38, and for ZmGLN1, between amino acids 43 and 44. However, analysis of the ZmGLN1 cryogenic electron microscopy (cryo-EM) structure showed that amino acids 31–43 are involved in decamer formation. For most proteins, the cTPs are not completely cleaved off; instead, a portion is retained to participate in the conformational formation of the mature protein43. Therefore, we propose that the cleavage site is located between amino acids 30 and 31 (Fig. 4a). To validate the above hypothesis, we performed truncation experiments and assessed protein subcellular localization using fluorescence markers. For ZmNIR2, deletion of the entire cTP (Δ47 (ZmNIR2)-eGFP) resulted in the protein failing to enter chloroplasts. Deletion of the first 37 amino acids (Δ37 (ZmNIR2)-eGFP) led to partial protein accumulation around the chloroplasts, probably because of recognition by translocon complexes on the chloroplast membrane. However, the protein still could not enter the chloroplasts (Fig. 4c). For ZmGLN1, deletion of the entire cTP (Δ51 (ZmGLN1)-eGFP) and deletion of the first 30 amino acids (Δ30 (ZmGLN1)-eGFP) both caused the protein to be unable to enter chloroplasts (Fig. 4d). Fluorescence localization results from truncation experiments support these predictions and further define the chloroplast-targeting mechanisms of ZmNIR2 and ZmGLN1.Fig. 4: ZmNIR2 and ZmGLN1 form a PG-dependent metabolic enzyme association.The alternative text for this image may have been generated using AI.Full size imagea,b, Protein segmentation diagram of maize ZmNIR2 and ZmGLN1 proteins. cTP, chloroplast transit peptide; cTP-CS, chloroplast transit peptide cleavage site; N/S-Fd-like, nitrite/sulfite reductase ferredoxin-like domain; N/S-4Fe-4S, nitrite/sulfite reductase 4Fe-4S; N-HR, ZmNIR2-hydrophobic-region; and G-HR, ZmGLN1-hydrophobic region. c,d, Representative fluorescence images of eGFP-tagged ZmNIR2 and ZmGLN1 variants in a and b, co-localized with PSY3-mCherry. Scale bar, 1 µm (partial images), 20 µm (complete images). e, The cryo-EM map of maize ZmGLN1. Top view (top, five-fold symmetry axis) and side view (bottom, two-fold symmetry axis). ZmGLN1 is a decameric structure composed of two identical pentameric rings stacked face to face. Each subunit is shown in a different colour. f, The structure model of maize ZmGLN1 in two view orientations, coloured as in e. g,h, Molecular dissection of the ZmGLN1 decamer. The structure is split into two pentameric rings (side views), each rotated 90° to show the inner and outer surfaces. i, Two pentamers interpenetrate and interlock by their N-terminal β-sheets, forming a β-barrel-like cylindrical structure. j, BN-PAGE analysis of the endogenous oligomerization of ZmGLN1 in the control (CK) and Zmgln1. k, BN-PAGE analysis of the endogenous oligomerization of ZmGLN1 in CK treated with 2 mM, 4 mM and 8 mM nitrogen concentrations. UGPase was used as the internal reference. l, Western blot analysis of the His semi-in vivo Co-IP assays with ZmNIR2T1OE plant lysates and ZmGLN1-His. Actin was used as the internal reference. m, Western blot analysis of the flag pull-down assays with ZmNIR2-flag and ZmGLN1-HA. An asterisk indicates impurity. j–m, The experiment was independently repeated three times with similar results. A representative result is shown. For gel source data, see Supplementary Fig. 4. n, FIDA binding analysis of ZmGLN1–ZmNIR2 interaction. Apparent dissociation constant (Kd) was determined by fitting concentration-dependent hydrodynamic radius changes of fluorescently labelled protein. Data are mean ± s.d.; n = 3 biologically replicates.Source dataAfter entering the chloroplast, ZmNIR2 and ZmGLN1 are further targeted to PGs by hydrophobic domains. Based on predicted transmembrane helix sequences with high hydrophobicity (Expasy-ProtScale, https://web.expasy.org/protscale/), we identified six consecutive (>10 aa) hydrophobic regions in ZmNIR2, designated as N-HR1 (aa 187–201), N-HR2 (aa 282–323), N-HR3 (aa 403–413), N-HR4 (aa 469–485), N-HR5 (aa 520–543) and N-HR6 (aa 566–597) (Fig. 4b and Extended Data Fig. 7a). We used truncation experiments to verify the PG-targeting signal regions of ZmNIR2. The results showed that removal of each of the six hydrophobic regions individually partially weakened the PG-targeting ability of ZmNIR2, increasing the amount of fluorescent protein retained in the chloroplasts (Extended Data Fig. 7b). Individual hydrophobic regions fused to the cTP of RuBisCO small subunit 1 (tpRBcS1) at their N-terminus failed to target PGs, even when they entered the chloroplasts (Extended Data Fig. 7c,d). ZmNIR2 completely lost its PG-targeting ability on removal of all six hydrophobic regions (Fig. 4c). These results demonstrate that the PG-targeting ability of ZmNIR2 relies on all six hydrophobic regions.ZmGLN1 contains two consecutive hydrophobic regions, designated as G-HR1 (aa 40–53) and G-HR2 (aa 393–423) (Fig. 4b and Extended Data Fig. 7e). ZmGLN1 retained partial PG-targeting ability on removal of G-HR1 (ΔG-HR1). However, deletion of G-HR1 impaired the chloroplast import, causing a portion of the protein to be retained in the cytoplasm. This hydrophobic region is proximal to the cTP, and its deletion might partially affect the folding state of the protein precursor or its interaction with cytosolic chaperones required for chloroplast preprotein import44. Removal of G-HR2 (ΔG-HR2) from ZmGLN1 completely abolished its PG-targeting ability (Extended Data Fig. 7f). Simultaneous removal of both G-HR1 and G-HR2 (ΔG-HR1-2) resulted in partial cytoplasmic retention of the protein, whereas the portion that entered the chloroplasts completely lost its PG-targeting ability (Fig. 4d). Individual G-HR1 and G-HR2 fragments lack a chloroplast transit peptide and thus could not enter chloroplasts (Extended Data Fig. 7g). When fused at their N-terminus to tpRBcS1 to facilitate chloroplast import45, RtpRbcS1-G-HR1 was not targeted to PGs, whereas tpRbcS1-G-HR2 exhibited partial PG-targeting ability (Extended Data Fig. 7h). These results indicate that the PG-targeting ability of ZmGLN1 depends on both G-HR1 and G-HR2, with G-HR2 playing a dominant part.ZmNIR2 and ZmGLN1 form a metabolonMaize possesses six ZmGLNs isoforms. However, as shown above, only ZmGLN1 (a plastidic GS2) is specifically localized to PGs within chloroplasts (Fig. 3e). To gain structural insights into the functional mechanism of ZmGLNl and its role in nitrogen assimilation, we performed high-resolution cryo-electron microscopy (cryo-EM) analysis of recombinant ZmGLN1 protein purified from Escherichia coli through a multi-step chromatography process, including affinity, ion exchange and size-exclusion chromatography. The cryo-EM structure of ZmGLN1 shows a decameric assembly with overall dimensions of 143.1 Å × 146.1 Å × 98.3 Å, exhibiting D5 symmetry characterized by five 2fold axes perpendicular to a central 5-fold axis (Fig. 4e-h, Extended Data Fig. 8, Supplementary Video 1 and Supplementary Table 1). This oligomeric state aligns with maize cytosolic ZmGS1a (ZmGLN5) but is distinct from the dodecameric architecture of cyanobacterial GS (Extended Data Fig. 9). This decameric structure is composed of two identical pentameric rings stacked face-to-face, resulting in 10 active sites, each formed at the interface between two neighbouring subunits within a ring. Two pentamers interpenetrate and interlock through their N-terminal β-sheets, resulting in a cylindrical structure with a β-barrel-like architecture (Fig. 4i). Comparison with the maize cytosolic ZmGS1a structure shows high conservation of residues involved in ADP and glutamate-analogue binding (Supplementary Fig. 3), suggesting that both the oligomeric assembly and catalytic mechanism of GLN1 are conserved with plant cytoplasmic GS1a.Using a ZmGLN1-specific endogenous antibody, we examined the native oligomeric state of ZmGLN1 in planta in both CK (control) and gln1 mutant maize. In CK, a single band near 440 kDa was observed, whereas it was absent in the Zmgln1 mutant. Based on the estimated molecular weight of the ZmGLN1 monomer (46 kDa), the result supports the decameric state of GLN1 in plants (Fig. 4j). Dynamic light scattering (DLS) analysis shows a dominant population of recombinant GLN1 in solution with hydrodynamic radii of 7.19 ± 0.07 nm and 7.17 ± 0.05 nm, corresponding to hydrodynamic diameters of approximately 14.3–14.4 nm and in agreement with the overall dimensions of the decameric assembly determined by cryo-EM (143.1 Å × 146.1 Å × 98.3 Å). Together, these results indicate that ZmGLN1 exists stably as a decamer in plants. To investigate how this oligomeric state changes under varying nitrogen conditions, we treated maize seedlings with three nitrogen concentrations (2 mM, 4 mM and 8 mM) following the cultivation method described (Fig. 1a). Equal amounts of leaf tissue were then used to assess ZmGLN1 oligomerization. Given that the number of PGs itself increases with rising nitrogen concentrations, we similarly observed an increase in ZmGLN1 decamers in response to higher nitrogen levels (Fig. 4k).Given the sequential catalysis of nitrite to glutamine by PG-localized NIR2 and GLN1, we asked whether they interact to enhance assimilation efficiency. To determine whether GLN1 is involved in the potential interaction with NIR2 in PGs, we performed unbiased immunoprecipitation-mass spectrometry (IP-MS) using transgenic plants overexpressing Flag-tagged NIR2. As expected, GLN1 was identified among the interacting proteins (see Source data). To validate the interaction between these key nitrogen assimilation enzymes, ZmNIR2 and ZmGLN1, we carried out a semi-in-vivo Co-IP assay. ∆30 (ZmGLN1)-His, purified from E. coli, was co-precipitated with ZmNIR2 from lysates of Flag-NIR2T1OE plants (Fig. 4l). Consistent with this, the in vitro pull-down assays confirmed that recombinant ∆30 (ZmNIR2)-Flag can pull down recombinant ∆30 (ZmGLN1)-HA (Fig. 4m). Furthermore, Flow-induced dispersion analysis (FIDA) quantified the binding affinity between recombinant ZmNIR2-Flag and ZmGLN1-HA, yielding an apparent binding constant (apparent Kd) of approximately 54 nM (Fig. 4n). Together, these results support a model in which ZmNIR2 and ZmGLN1 form a metabolon46,47 in PGs that spatially couples nitrite reduction and ammonium assimilation, thereby enhancing the efficiency of nitrogen flux from NO2− to NH4 and ultimately to glutamine.
Plastoglobules compartmentalize nitrogen assimilation in maize - Nature
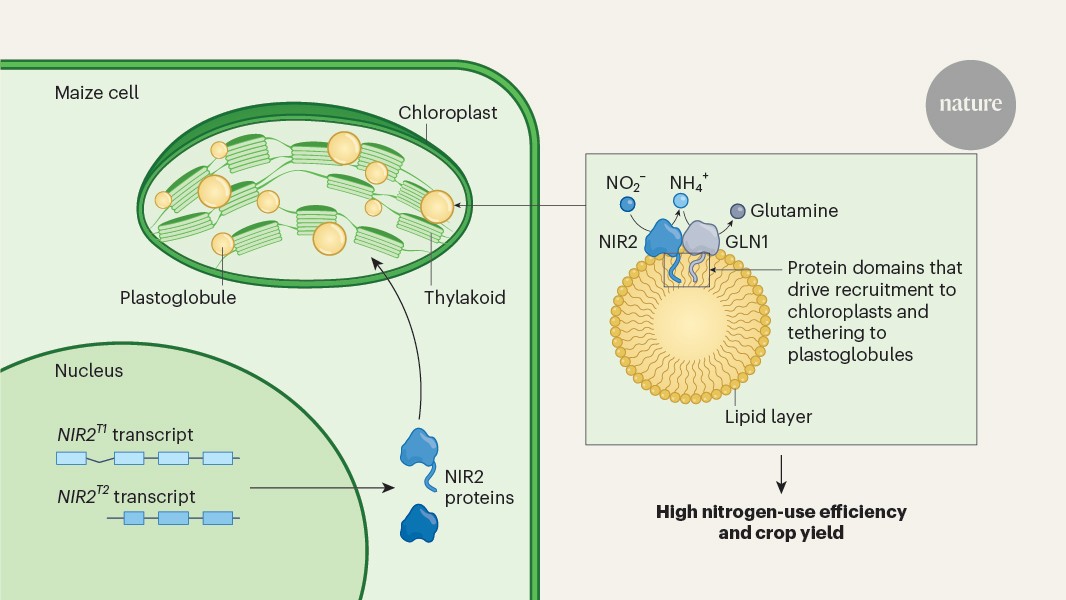
Chloroplast plastoglobules act as nitrogen-assimilation hubs in maize, with key enzymes enhancing nitrogen-use efficiency and offering new strategies for developing high-yield, sustainable crops.
14,509 words~66 min read