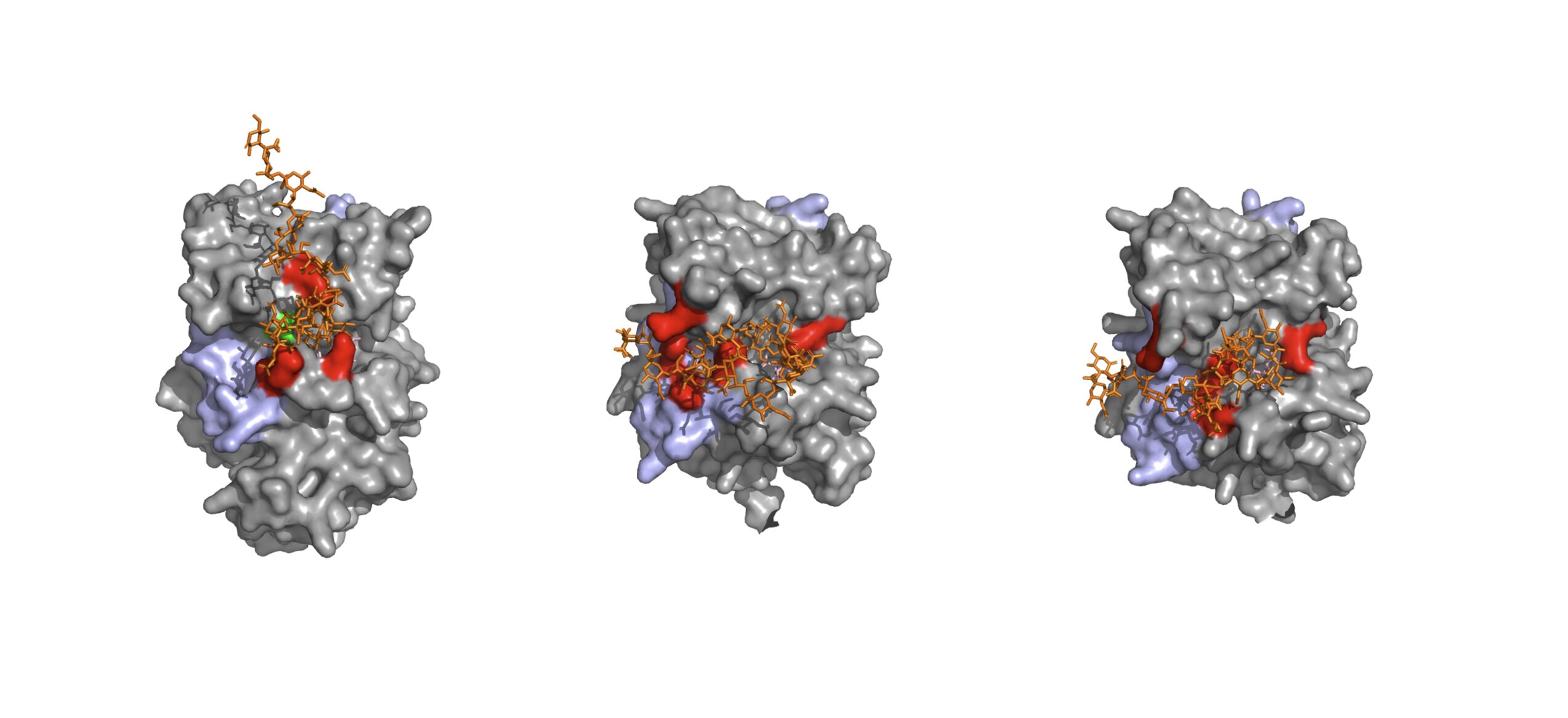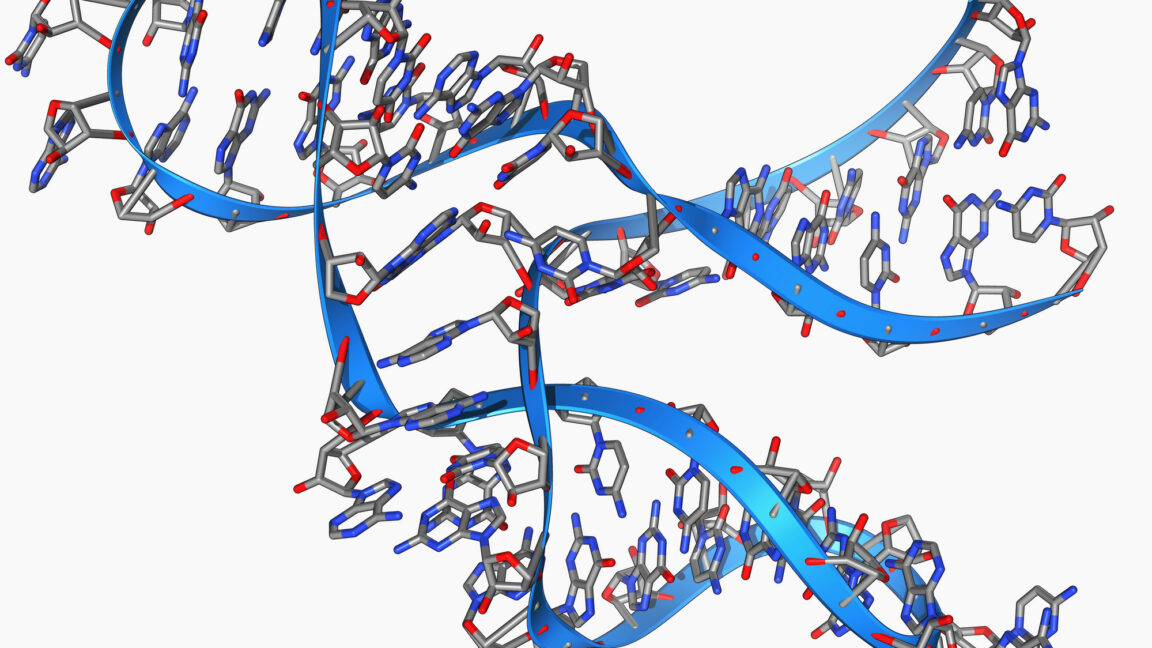
MainRNA polymerase II (RNAPII) extensively transcribes mammalian genomes, yielding a wide range of unadenylated and polyadenylated RNAs1,2,3,4. Moreover, individual transcription units that generate standard full-length transcripts also give rise to an array of shorter isoforms9,10. Thus, functional RNAs are produced alongside a wealth of futile RNAPII products. Whereas mature functional pA+ RNAs, such as protein-coding mRNAs, are exported from the nucleus to the cytoplasm, their non-functional counterparts are typically retained and degraded3,4. This is primarily achieved by the nucleoplasmic PAXT connection, which consists of a heterodimeric core of the RNA helicase MTR4 and the Zn-finger protein ZFC3H18,11. Additional, and less well-described, interactions with the nuclear pA+ RNA-binding protein PABPN1 and other transiently interacting RNP components, sometimes referred to as extended PAXT components, may aid in directing transcript turnover by the 3′−5′ exonucleolytic exosome complex12,13,14,15. However, how these interactions provide a biochemical basis by which PAXT distinguishes non-functional pA+ RNAs remains a major unresolved question.Prior to their nuclear export, pA+ RNAs are packaged with proteins into pA+ RNPs. Central to this process is the export factor and DExD-box ATPase UAP56, which is recruited to pA+ RNPs in preparation for their nuclear export5,16. At the nuclear envelope, the activity of the nuclear pore complex (NPC)-associated GANP–PCID2–SEM1 (GANP–PS) trimer of TREX-26 facilitates the release of RNA from UAP56, enabling export7,17. Again, how pA+ RNP sorting is orchestrated to favour the selected export of functional pA+ RNAs is unknown.Here we interrogate two TREX-2-like human complexes, SAC3D1–PCID2–SEM1 (SAC3D1–PS) and LENG8–PCID2–SEM1 (LENG8–PS), in which the conserved SAC3D1 and LENG8 proteins, respectively, replace GANP. The GANP–PS, SAC3D1–PS and LENG8–PS complexes are structurally similar and share the ability to release UAP56 from RNA. Notably, we show that LENG8–PS offers PAXT a module, that acts on UAP56 to promote transcript turnover in contrast to the RNA export activity of TREX-2. Our findings reveal that nuclear pA+ RNA export and decay utilize a shared biochemical mechanism to act on pA+ RNPs but with fundamentally different outcomes. Based on the substrate preference of PAXT and its separate nuclear localization from TREX-2, we propose a general model for pA+ RNP fate determination.TREX-2-like complexes release RNA from UAP56To mediate the docking of export-competent pA+ RNPs at the NPC, UAP56 binds the five subunit TREX-2 complex7 (GANP, PCID2, SEM1, CETN2 or CETN3, and ENY2). Within this complex, UAP56 contacts the TREX-2 complex core (TREX-2M), which comprises PCID2, SEM1 and the SAC3 domain of the scaffolding subunit GANP7 (Fig. 1a, left). The ability of TREX-2M to release UAP56 from the pA+ RNP depends on the conserved ‘wedge loop’ within the SAC3 domain7,17 (Fig. 1b). Notably, similar SAC3 domains are found in the UAP56-interacting LENG8 and SAC3D1 proteins7. Although they are broadly conserved amongst eukaryotes, these proteins share no sequence features with GANP or each other aside from the SAC3 domain (Fig. 1a,b). Moreover, proteome-wide AlphaFold2 screens suggested that SAC3 domains of LENG8 or SAC3D1 form complexes with PCID2 and SEM118,19,20, thus mimicking TREX-2M. Finally, and central to the present study, LENG8 co-immunoprecipitated with PAXT core components ZFC3H1 and MTR48,21 (also see Fig. 2 below) and was shown to interact with PCID2 and SEM1 in both human and yeast cells22,23. Collectively, this prompted us to investigate these TREX-2M-like complexes in more detail.Fig. 1: TREX-2-like complexes bind UAP56 to trigger RNA release.a, Cartoons of TREX-2 and TREX-2-like core complexes (top) and their domain architectures (bottom). PCID2, dark blue; SEM1, mid blue; SAC3 domain-containing proteins GANP, LENG8 and SAC3D1, shades of blue. Wedge loop positions are shown as grey bars. Regions included in the atomic models in e,f are indicated by black lines. WH, winged helix. b, Multiple sequence alignment of wedge loop sequences of human GANP (UniProt O60318), LENG8 (UniProt Q96PV6) and SAC3D1 (UniProt A6NKF1), with a conserved tyrosine residue anchoring the wedge loop on the SAC3 domain (Yanchor) and the central wedge loop arginine (Rwedge) highlighted. Colouring by conservation (blue letters, conserved residue; blue background, invariant residue). c,d, UAP56 RNA release assay, demonstrating the stimulatory effects of LENG8–PSM (c) or SAC3D1–PSM (d) complexes. Bead-immobilized 15 poly-uridine RNA was incubated with UAP56 and ATP to form UAP56–ADP-Pi–RNA complexes7 and subsequently challenged with recombinant LENG8–PSM or SAC3D1–PSM complexes, or their respective wedge loop mutants. Remaining UAP56–ADP-Pi-RNA complexes were analysed by SDS–PAGE and Coomassie staining. e,f, Cartoon representation of cryo-EM structures of UAP56–RNA–LENG8–PSM (e) or UAP56–RNA–SAC3D1–PSM (f) complexes at resolutions ranging from 6 to 12 Å for UAP56–RNA–LENG8–PSM and 2.6 to 4.5 Å for UAP56–RNA–SAC3D1–PSM. SEM1, blue; PCID2, dark blue; LENG8491–800, light blue; SAC3D148–404, green blue; UAP56, shades of pink; flexible region of UAP56 N-terminal domain, dashed line; RNA, black. e, Bottom left, UAP56 domain structure. g, Details of the SAC3D1 wedge loop–UAP56–nucleotide interactions for UAP56–RNA–SAC3D1–PSM (left) or UAP56–RNA–TREX-2M (right) (stick representation). Superpositions of the wedge loop anchoring tyrosine and the central arginine of SAC3D1 or GANP are shown in the middle. GANP, light blue; other colours as in a. h, Cartoon model of UAP56–RNA–TREX-2 or TREX-2-like complex interactions, including RNA release from UAP56. Left to right: (1) pre-RNA release state: TREX-2 and TREX-2-like complexes bind RNA-clamped UAP56; (2) post-RNA release state: RNA is unclamped from UAP56, leaving UAP56 in an open conformation bound to the TREX-2 or TREX-2-like complex; (3) dissociation.Fig. 2: LENG8–PS constitutes a TREX-2-like module of PAXT.a, Immunofluorescence analyses of central TREX-2 and TREX-2-like components. Anti-Flag antibody (left column)- and DAPI (mid column)-stained HeLa cell lines (merged signals, right column), expressing C-terminally 3×Flag-tagged endogenous GANP (top row), LENG8 (second row), SAC3D1 (third row) or PCID2 (bottom row). Scale bars, 10 μm. b, Volcano plots of Flag IP–MS analyses of extracts from 3×Flag-tagged GANP (left), LENG8 (mid) or PCID2 (right) cells from a. log2 fold label-free quantification (LFQ) changes of interactor signals in the individual immunoprecipitations over their maternal HeLa cell line control were plotted against −log10-transformed two-sided limma-moderated Student’s t-test P values calculated over biological triplicate data. TREX-2-like, core PAXT, exosome, TREX-2 and NPC components are colour-coded and labelled. c, Heat map of mean intensity-based absolute quantification (iBAQ) values, with control immunoprecipitation values subtracted, from triplicate immunoprecipitation experiments with 3×Flag-tagged GANP, LENG8 and PCID2, conducted without (−) or with (+) Pierce universal nuclease treatment. Displayed proteins as in b, and with EXOSC1–EXOSC5 and EXOSC10. d, Colocalization coefficients between ZFC3H1 and Flag immunofluorescence signals in maternal HeLa cells (from Extended Data Fig. 4j) and HeLa cells expressing 3×Flag-tagged ZFC3H1, LENG8, PCID2 or GANP. Red (displayed in magenta) and green channels were used for the detection of ZFC3H1 and Flag signals, respectively. Example cells with staining overlap and total numbers (n) of cells imaged are indicated above the plot. In all box plots, the centre line is the median, box edges delineate the interquartile range and whiskers represent the distribution of data points within 1.5× interquartile range (Methods). e, AlphaFold2 model of interacting regions of ZFC3H1 (top) and LENG8 (bottom), shown in cartoon representation with interface residue as sticks (middle). The conserved F301 of LENG8, which is critical for interaction, is highlighted. f, Volcano plot as in b, but displaying ZFC3HΔ730−747–3×Flag relative to wild-type ZFC3H1–3×Flag sample data. Constructs were expressed in HeLa cells expressing ZFC3H1–2×HA–dTAG that were treated with dTAGV-1 to deplete endogenous ZFC3H1. Note additional colour coding of extended PAXT components and UAP56 (see g). WT, wild type. g, As in f, but for LENG8(F301A)–3×Flag relative to wild-type LENG8–3×Flag constructs expressed in dTAGV-1-treated cells expressing LENG8–2×HA–dTAG, h. Cartoon depicting localization-distinct TREX-2-like modules.Given the critical role of UAP56 in pA+ RNP export via TREX-2, we hypothesized that LENG8 and SAC3D1 might target UAP56-bound RNPs to different cellular fates. To address this, we first explored the structure–function relationships of LENG8 or SAC3D1 with UAP56 in vitro. As previously achieved for GANP7, we purified stable recombinant complexes of the SAC3 domain-containing constructs of LENG8491–800 or SAC3D148–404 in the presence of PCID2–SEM1 (constituting LENG8–PSM or SAC3D1–PSM; Supplementary Fig. 2a,b). Both complexes could bind UAP56 in the presence of the non-hydrolysable ATP analogue adenylyl-imidodiphosphate (AMP-PNP) and a 15-nucleotide poly-U RNA substrate (Extended Data Fig. 1a,b, lanes 1–5). TREX-2M facilitates the release of ADP and Pi from UAP56, thus accelerating the rate-limiting step in the disassembly of UAP56–RNA complexes, releasing free UAP56 available for RNA re-binding, and resulting in an increased apparent ATPase activity7. Similarly, the LENG8–PSM or SAC3D1–PSM complexes stimulated the apparent ATPase rate of UAP56 in the presence of RNA and ATP in vitro, revealing approximately a 290-fold and 60-fold stimulation, respectively (Extended Data Fig. 1c,d). Moreover, substituting a highly conserved arginine residue in the LENG8 or SAC3D1 wedge loops with an alanine7 (LENG8(R563A) or SAC3D1(R102A)) (Fig. 1b, Extended Data Fig. 1e and Supplementary Fig. 2a,b), did not affect UAP56 binding (Extended Data Fig. 1f,g and Supplementary Fig. 2c), but largely abrogated the ATPase stimulatory activity on UAP56 (Extended Data Fig. 1h,i). Finally, to test whether LENG8–PSM and SAC3D1–PSM, like TREX-2M (ref. 7), would promote the release of RNA from UAP56, we incubated UAP56 with RNA and ATP to form UAP56–ADP-Pi–RNA complexes, which we immobilized on streptavidin beads via the biotinylated RNA7. These complexes were then challenged with either LENG8–PSM or SAC3D1–PSM, revealing that both moieties released UAP56 efficiently (Fig. 1c,d, compare lanes 4 and 5), whereas the respective wedge loop mutants did not (Fig. 1c,d, lane 6). Of note, mutating three residues targeting the UAP56 N-terminal domain (NTD) and UAP56 RecA2-binding interfaces of LENG8 (LENG8(TRR)) led to diminished LENG8–PSM–UAP56 interaction (Extended Data Fig. 1j,k and Supplementary Fig. 2a) and parallel declines in both the apparent ATPase activity (Extended Data Fig. 1h, lane 7) and the release of UAP56 from RNA (Extended Data Fig. 1l). We conclude that TREX-2-like complexes, like TREX-2, bind UAP56 and trigger the release of its bound RNA through a shared mechanism.Although previous structural studies of UAP56–TREX-2M complexes had revealed their protein–protein interfaces, it remained unclear how the wedge loop functions in releasing UAP56 from RNA. To investigate the molecular basis for this function, we analysed LENG8–PSM and SAC3D1–PSM complexes with UAP56 in the presence of 15-nucleotide poly(U) RNA and ATP or AMP-PNP using cryo-electron microscopy (cryo-EM). This revealed a fraction of complexes without UAP56, enabling us to solve the structures of apo LENG8–PSM and SAC3D1–PSM at 3.5 Å and 3.6 Å resolution, respectively (Extended Data Table 1). Both complexes showed the same V-shaped architecture previously observed for TREX-2M (Extended Data Figs. 2a and 3 and Supplementary Figs. 3a–c and 4a–c) and a yeast LENG8–PSM complex23,24. Unexpectedly, two-dimensional class averages of the UAP56-engaged fractions of LENG8–PSM and SAC3D1–PSM suggested that UAP56 could be in a closed, RNA-bound state, prior to its release via the wedge loop (Extended Data Fig. 2b). Together with our previously reported UAP56–TREX-2M structure7, in which UAP56 was captured after RNA release, this enabled us to investigate the RNA-releasing mechanism of SAC3 domain-containing complexes. We resolved the cryo-EM structures of UAP56–LENG8–PSM and UAP56–SAC3D1–PSM complexes in the pre-RNA release state (Fig. 1e,f, Extended Data Fig. 3, Supplementary Figs. 3d–g and 4f,g and Extended Data Table 1). A severe bias in particle orientation limited resolution to 6–12 Å for UAP56–LENG8–PSM in the RNA-clamped pre-release state. Reconstitution of the complex with ATP yielded a higher resolution structure at 4.9 Å containing density only for the UAP56 NTD (Supplementary Fig. 4d,e). We could, however, resolve the structure of UAP56–RNA–SAC3D1–PSM to 2.6 Å, enabling a detailed structural analysis. The structure of the pre-release state shared key architectural features with UAP56–TREX-2M, including the anchoring of the NTD of UAP56 at the base of the SAC3D1–PS complex. Truncating the UAP56 NTD reduced the affinity of UAP56 for both SAC3D1–PSM and LENG8–PSM by more than 30-fold, as measured by grating-coupled interferometry (Extended Data Fig. 2c) and supported by in vitro pulldown assays (Extended Data Fig. 1a,b, lanes 6 and 7). Thus, the UAP56 NTD is equally important for TREX-2-like complex and TREX-2-complex7 interactions. In addition, the UAP56–SAC3D1–PSM structure provided insights into the action of the wedge loop. In the structure, this region (residues Y100–P111; Fig. 1b and Extended Data Fig. 1e) is bound near the two RecA lobes through largely electrostatic interactions between the peptide backbone and UAP56 residues R135 in RecA1 and K334 in RecA2 (Fig. 1g and Extended Data Fig. 2d). The critical R102 wedge loop residue in SAC3D1 forms a hydrogen bond with UAP56 E354, positioning R102 close to F381 in the RecA2 lobe of UAP56. By contrast, in the post-release state observed for UAP56–TREX-2M, this central wedge loop arginine (R102 in SAC3D1, R678 in GANP) replaced UAP56 F381 in the nucleotide binding site (Fig. 1g, right). The positioning of the wedge loop arginine in the clamped state might prime it to replace UAP56 F381 in a subsequent step, releasing RNA from UAP56 (Fig. 1g,h and ref. 7).The RNA-clamped RecA lobes of UAP56 are bound between PCID2, the wedge loop and the SAC3 domain in these SAC3 domain-containing complexes. Notably, the protein–protein interfaces between UAP56 and PCID2 in both TREX-2M and the TREX-2M-like complexes involve only few specific interactions, except for the UAP56 NTD7, suggesting that PCID2–SEM1 has an architectural role in ensuring specificity for UAP56. Indeed, superposition of the evolutionarily related and RNA-bound form of the DExD-box ATPase EIF4A316 onto the UAP56–SAC3D1–PSM structure revealed clashes between EIF4A3 and PCID2 (Extended Data Fig. 2e). Consistently, LENG8–PSM bound UAP56, but not the closely related DExD-box proteins EIF4A3 and DDX19 in vitro (Extended Data Fig. 2f and Supplementary Fig. 2d) and did not stimulate the EIF4A3 ATPase (Extended Data Fig. 2g).We conclude that human cells contain three structurally and biochemically equivalent SAC3 domain-containing complexes. Aided by their complex architecture and the uniqueness of the UAP56 NTD, they all target UAP56 specifically and their conserved wedge loops can dislodge UAP56 from RNA.LENG8–PS provides a physical module for PAXTOur biochemical and structural analyses suggested that the GANP–PS, LENG8–PS and SAC3D1–PS complexes can individually act on UAP56. To address where these complexes act in vivo, we generated HeLa cell lines25 stably expressing C-terminally 3×Flag-tagged versions of endogenous GANP, LENG8 or SAC3D1 as well as the common subunit PCID2 (Extended Data Fig. 4a, lanes 3–6), and analysed these with immunofluorescence microscopy using a Flag antibody. As previously reported, GANP was found primarily at the nuclear envelope, consistent with its NPC association6,26 (Fig. 2a, top row). By contrast, LENG8 and SAC3D1 localized to the nucleoplasm and the cytoplasm, respectively (Fig. 2a, second and third rows). In agreement with its presumed presence in all three complexes, PCID2 was distributed between the nucleoplasm, the nuclear envelope and the cytoplasm (Fig. 2a, bottom row).With our focus on nuclear RNA sorting, we examined the TREX-2 and LENG8–PS complexes in more detail, performing immunoprecipitation–mass spectrometry (IP–MS) analyses of the 3×Flag-tagged GANP, LENG8 and PCID2 proteins. Stringent immunoprecipitation conditions were used to enrich for high-affinity interactors (Methods). In the GANP–3×Flag immunoprecipitation, this yielded large amounts of PCID2, in addition to the ENY2 and CETN2 or CETN3 subunits of TREX-26, and the nuclear pore basket protein TPR26 (Fig. 2b, left). Although LENG8–3×Flag also precipitated PCID2, this immunoprecipitation was instead enriched for the PAXT core components ZFC3H1 and MTR4 along with nuclear exosome subunits (Fig. 2b, middle; note that SEM1 was also detected applying an alternative protocol (Extended Data Fig. 5l)). Finally, reflecting its dual interaction with GANP and LENG8, PCID2–3×Flag precipitated these proteins and their respective interactors (Fig. 2b, right). Parallel, low-stringency immunoprecipitation–western blotting analyses qualitatively recapitulated these interaction patterns (Extended Data Fig. 4a, lanes 9–12) and subjecting the IP–MS experiments to RNase treatment revealed that the interactions were not facilitated by RNA (Extended Data Fig. 4b). Moreover, analysing mean enrichments over background across immunoprecipitation experiments displayed near-identical interaction levels of GANP with TREX-2 components, and LENG8 precipitated similar amounts of PCID2 as well as the core PAXT factors ZFC3H1 and MTR4 (Fig. 2c and Supplementary Table 1). This indicated that LENG8–PS complexes constitute a major module for PAXT. To address this notion further, we generated ZFC3H1–3×Flag cells (Extended Data Fig. 4a, lane 2) and conducted IP–MS experiments at both low and high stringency, which revealed RNase-resistant interactions with LENG8 and PCID2 (Extended Data Fig. 4c,d). Mean enrichments calculations showed that, along with exosome subunits, LENG8 and PCID2 were sub-abundant to MTR4, consistent with the reported presence of inactive nuclear ZFC3H1–MTR4 dimers12 (Extended Data Fig. 4e, columns 1 and 2). Nonetheless, the ZFC3H1 immunoprecipitations returned LENG8 and PCID2 in 5- to 10-fold excess of extended PAXT components and reciprocal LENG8 immunoprecipitations recovered abundant amounts of the ZFC3H1–MTR4 dimer (Extended Data Fig. 4e, columns 3 and 4, f). We further reproduced key high-stringency interactions in HCT116 cells, expressing endogenous LENG8–2×HA–dTAG or ZFC3H1–2×HA–dTAG proteins (Extended Data Fig. 4g–i and Supplementary Table 1). Finally, association of LENG8 with ZFC3H1 was mirrored by their measured nuclear colocalization, as revealed by immunostaining of LENG8–3×Flag cells using Flag- and ZFC3H1-specific antibodies. Here, LENG8 and PCID2, but not GANP, displayed weighed colocalization coefficients with ZFC3H1 in the nucleoplasm of more than 0.90 (Fig. 2d and Extended Data Fig. 4j).Having established a physical link between LENG8 and ZFC3H1, we inquired whether LENG8 interacts with MTR4 and the exosome via ZFC3H1. Indeed, rapid ZFC3H1 depletion, using the FKBP12(F36V)–degron (dTAG)27 (Extended Data Fig. 5a, left), prevented interactions of MTR4 and the exosome component EXOSC10 with LENG8–3×Flag (Extended Data Fig. 5a, right). To identify a possible ZFC3H1 interaction site on LENG8, we utilized AlphaFold2, which revealed a conserved motif of two α-helices (residues 288–342) in the otherwise unstructured region N-terminal to the SAC3 domain (Extended Data Fig. 5b–d). A direct interaction was predicted between this helical region of LENG8 and a single α-helix (residues 730–747) of ZFC3H1 (Fig. 2e and Extended Data Fig. 5e), which we verified using an in vitro pulldown assay, comparing relevant wild-type or mutant versions of recombinant LENG8 and ZFC3H1 peptides (Extended Data Fig. 5f and Supplementary Fig. 2e). Further validating the interaction, IP–MS analysis of a ZFC3H1(Δ730–747) compared to a wild-type ZFC3H1 3×Flag construct demonstrated the specific loss of LENG8 and PCID2 over exosome subunits and extended PAXT components (Fig. 2f and Extended Data Fig. 5g) also upon nuclease treatment (Extended Data Fig. 5h). Similarly, mutation of the central LENG8 phenylalanine at the LENG8–ZFC3H1 interface (Fig. 2e) to an alanine (F301A) led to the selective loss of ZFC3H1, MTR4 and exosome subunits in a comparative IP–MS analysis of wild-type LENG8 versus LENG8(F301A) 3×Flag constructs (Fig. 2g, Extended Data Fig. 5i–l and Supplementary Table 1).Together, we conclude that the TREX-2-like LENG8–PS complex constitutes a physical module of the nucleoplasmic PAXT connection (Fig. 2h).LENG8–PS works with PAXTGiven the physical link between ZFC3H1 and LENG8, we next sought to probe its functional relevance. Expression of wild-type ZFC3H1–3×Flag, but not the ZFC3H1(Δ730–747) LENG8 binding-deficient variant (Extended Data Fig. 6a), suppressed selected formerly established PAXT substrates8,14 from accumulating after rapid depletion of endogenous ZFC3H1 (Fig. 3a). Similarly, PAXT substrate accumulation, following rapid depletion of endogenous LENG8, was suppressed by expression of wild-type LENG8–3×Flag but not by the ZFC3H1-binding mutant LENG8(F301A) (Fig. 3b and Extended Data Fig. 6b). Since these analyses corroborated a role for LENG8 in PAXT function, we obtained a transcriptome-wide view of this relationship by sequencing pA+ RNA from cells rapidly depleted for either LENG8 or ZFC3H1 (Extended Data Fig. 6c,d). PAXT substrates include numerous prematurely terminated transcripts (PTTs), deriving from transcription start site (TSS)-proximal regions of protein-coding genes8,14. To incorporate these in our analysis, we identified transcription units displaying such increased pA+ RNA coverage upon depletion of ZFC3H1 or LENG8 (Extended Data Fig. 6e) and intersected data with pA+ RNA 3′ end peaks previously identified upon depletion of ZFC3H128 (Methods). As a result, 1,202 pA+ PTTs were defined (see Extended Data Fig. 6f for an example) and included in our HeLa transcriptome annotation29 (Extended Data Fig. 6g).Fig. 3: LENG8–PS provides a functional module for PAXT.a, Box plots of RT–qPCR analysis of selected PAXT substrates from cells expressing ZFC3H1–2×HA–dTAG and treated with (+) or without (−) dTAGV-1 for 4 h and complemented by expression of wild-type ZFC3H1 or ZFC3H1(Δ730–747) as indicated. The prefixes pt and pro indicate prematurely terminated and PROMPT, respectively. Values are relative to mock-treated HeLa cells and normalized to GAPDH RNA expression. Coloured dots denote the mean value of n = 3 biological replicates. Two-sided Bonferroni-corrected Student’s t-tests, calculated between conditions and combining the results for all RNAs tested. Similar statistical tests were conducted for the subsequent RT–qPCR analyses. b, As in a but for cells expressing LENG8–2×HA–dTAG and treated with (+) or without (−) dTAGV-1 for 24 h and complemented by expression of wild-type LENG8 or LENG8(F301A) as indicated. c, Scatter plot of mean log2 fold abundance changes in pA+ RNA of n = 3 biological replicates after treatment of cells expressing ZFC3H1–2×HA–dTAG (y axis) or LENG8–2×HA–dTAG (x axis) with 500 nM dTAGV-1 for 4 h. The trendline is shown in red and Pearson correlation coefficient (r) is indicated. d, Counts of differentially expressed pA+ RNAs after treatment of cells expressing ZFC3H1–2×HA–dTAG or LENG8–2×HA–dTAG with dTAGV-1 for 4 h. Transcripts are colour-coded by the number of exons. DESeq2 (adjusted P value < 0.1) was used and RNAs with log2 (fold change) greater than 0.5 and less than −0.5 were counted as upregulated and downregulated, respectively, here and in subsequent analyses. e, Box plots of log2 fold abundance changes in pA+ RNA from c after dTAGV-1 treatment of cells expressing ZFC3H1–2×HA–dTAG (top) or LENG8–2×HA–dTAG (bottom). All RNAs with measurable fold changes (P value < 0.1, per DESeq2) in either of the depletions were grouped by their contained exon numbers. Numbers of RNAs (n) examined are shown below the plots. f, Box plots of log2-scaled means of n = 3 biological RNA-seq replicates of nuclear/cytoplasmic ratios in unperturbed HeLa cells of upregulated ZFC3H1- or LENG8-sensitive pA+ RNAs compared to all other annotated RNAs. Two-sided unpaired Welch’s t-test. g, Counts of all annotated (left) or PAXT-sensitive (right) mRNAs stratified by contained exon numbers. PAXT sensitivity was scored as upregulation upon ZFC3H1 or LENG8 depletion. mRNAs containing 1 to 4 exons (classified as low-exon mRNAs) are marked with a pink bar.Although numbers and changed levels of affected RNAs were higher with ZFC3H1 depletion, individual LENG8- and ZFC3H1-sensitive transcripts were strongly correlated (Fig. 3c), consistent with a shared pathway. This was substantiated by sequencing pA+ RNA from ZFC3H1-depleted cells, exogenously expressing either wild-type ZFC3H1 or the LENG8-binding mutant ZFC3H1(Δ730–747) (Extended Data Fig. 6h); only wild-type ZFC3H1 efficiently suppressed the accumulation of ZFC3H1 substrates, whereas ZFC3H1(Δ730–747) retained only partial activity (Extended Data Fig. 6i). Moreover, differential expression analysis, applying DESeq2 (ref. 30) with two different cut-offs, revealed an extensive overlap of RNAs that were increased upon depletion of ZFC3H1 or LENG8 (Extended Data Fig. 6j and Supplementary Table 2). We speculated that the stronger effect size with ZFC3H1 depletion might, at least in part, reflect the interaction of ZFC3H1 with the early RNA-processing factor ARS214, possibly allowing residual LENG8-independent ZFC3H1 activity. To test this, we compared ARS2- and LENG8-dependent PAXT targets by intersecting RNA-sequencing (RNA-seq) data from a ZFC3H1 complementation assay using the ZFC3H1(ARM) mutant, which cannot bind ARS214, with RNA-seq data from the LENG8 binding-deficient ZFC3H1(Δ730–747) mutant. There was significant overlap between LENG8- and ARS2-dependent ZFC3H1 substrates, which suggested partially redundant LENG8 and ARS2 activities on a shared substrate pool (Extended Data Fig. 6k).In agreement with previous studies on PAXT8,11,13,14, the ZFC3H1- and LENG8-depleted samples revealed upregulation of promoter upstream transcripts (PROMPTs)31, PTTs, other noncoding RNAs (ncRNAs) and a minor fraction of full-length mRNAs, both when each sample was evaluated individually (Extended Data Fig. 6l) and when interrogating the common substrate cohort (Extended Data Fig. 6m, left). In line with this result, the majority of ZFC3H1 and LENG8 substrates contained only one or a few exons (Fig. 3d and Extended Data Fig. 6m, right), and for both depletion conditions RNA accumulation levels decreased with increasing exon number (Fig. 3e and Extended Data Fig. 6n) and mature RNA length (Extended Data Fig. 6o,p).In addition to decay, ZFC3H1 also contributes to nuclear retention of pA+ RNA11,12,15, and LENG8 has been suggested to have a similar capacity32. To analyse the effect of ZFC3H1 or LENG8 on transcript fate, we therefore performed nuclear/cytoplasmic pA+ RNA-seq obtained by fractionating HeLa cells before or after the rapid depletion of these factors (Extended Data Fig. 7a,b). Consistent with the RNA-retention capacities of ZFC3H1 and LENG8, PAXT-sensitive RNAs, defined by their increased abundance upon either ZFC3H1 or LENG8 depletion (Methods), displayed higher nuclear-to-cytoplasmic ratios in unperturbed HeLa cells than the remaining pA+ transcriptome (Fig. 3f). Clustering all transcripts by their depletion-dependent nuclear or cytoplasmic content changes demonstrated that more than half of the displayed RNAs were immune to ZFC3H1 or LENG8 depletion (Extended Data Fig. 7c, left, cluster 1, Supplementary Table 2 and Methods). Whereas most of these transcripts were accounted for by spliced mRNAs with multiple exons (Extended Data Fig. 7c, biotypes), a closer examination of the low-exon count RNAs from cluster 1 revealed mild, but detectable, sensitivity to ZFC3H1 depletion (Extended Data Fig. 7d). Outside of cluster 1, the remaining transcripts were, to variable extents, upregulated in both compartments upon ZFC3H1 and LENG8 depletions, demonstrating inefficient nuclear retention (Extended Data Fig. 7a, lanes 1–4 and 6–9 and Extended Data Fig. 7c, clusters 2–4). Repeating the same depletion experiments in HCT116 cells (Extended Data Fig. 7e,f) recapitulated these trends (Extended Data Fig. 7g). It therefore appears that PAXT generally retains and mediates decay of short pA+ RNAs with few exons, whereas transcripts that escape these fates are commonly longer and more exon-rich.Although the protein-coding fraction of analysed transcripts was largely insensitive to PAXT, a minor subset of sensitive mRNAs was still detectable (Extended Data Fig. 7c, note biotypes of clusters 2–4). Similar to noncoding PAXT substrates, these were primarily low-exon (1–4 exons) transcripts (Fig. 3g), enriched in nuclei of unperturbed cells (Extended Data Fig. 7h). However, in the same condition, these short mRNAs were present at higher levels than their ncRNA counterparts (Extended Data Fig. 7i), suggesting that they might have acquired means to fend off nuclear turnover (see Discussion). DESeq2 also identified 105 longer, multi-exonic mRNA outliers (more than 4 exons), that were PAXT-sensitive despite their higher exon number and length. As general PAXT targets (Fig. 3f), these transcripts showed increased nuclear-to-cytoplasmic ratios in unperturbed cells compared with PAXT-insensitive controls (Extended Data Fig. 7j), suggesting that prolonged nuclear residence time may drive their sensitivity to PAXT. Notably, the LENG8 mRNA belongs to this transcript category, implying autoregulation of the PAXT pathway (see below). Since introns contribute to nuclear RNA retention33,34,35, we assessed PAXT-sensitive multi-exonic mRNAs (more than 4 exons) for reads spanning both 5′ and 3′ splice junctions (Methods). Indeed, when compared with a control population, these transcripts were enriched for retained introns, including previously described detained introns36 (Extended Data Fig. 7k). Moreover, upon ZFC3H1 or LENG8 depletion, PAXT-sensitive multi-exonic transcripts accumulated largely as incompletely spliced precursors in the nucleus and as fully spliced mRNAs in the cytoplasm (Extended Data Fig. 7l). Thus, upon PAXT impairment, this group of transcripts can be post-transcriptionally spliced and exported. Finally, we also identified an additional 3,209 PAXT-sensitive introns, including 161 detained introns (Supplementary Table 3), for which the corresponding spliced mRNAs were PAXT-insensitive as measured by DESeq2. We therefore propose that, in addition to short RNAs, PAXT can also target multi-exonic transcripts, which are retained in the nucleus due to incomplete splicing. In certain cases, this alters the cellular levels of the mature transcript, the extent of which might differ between cell types.Together, these cellular data indicate that LENG8–PS constitutes a functional module of PAXT, which primarily targets nuclear pA+ RNAs with no, or only a few, introns as well as a minor pool of longer intron-containing transcripts. This overall retention–decay regime may be exploited for the regulation of selected mRNAs.PAXT and TREX-2 govern UAP56-bound RNP fateHaving defined LENG8–PS as a central physical and functional module of PAXT, we investigated how the ability of LENG8 to release UAP56 from RNA in vitro (Fig. 1c and Extended Data Fig. 1l) affects its function in vivo. Similar to the ZFC3H1-binding–defective LENG8(F301A) mutant, the LENG8(TRR) and LENG8(R563A) variants (Extended Data Fig. 8a)—which were compromised for UAP56 binding (Extended Data Fig. 1j,k) and UAP56 release from RNA (Fig. 1c), respectively, in vitro—were unable to suppress selected PAXT targets that were upregulated upon depletion of endogenous LENG8 (Fig. 4a). Notably, compared with the wild-type and F301A constructs, LENG8(R563A) displayed reduced RNA binding in vivo (Extended Data Fig. 8b), despite its ability to bind major interactors (Extended Data Fig. 8c and Supplementary Table 1). We therefore reasoned that release of RNA from UAP56 by LENG8–PS is central for PAXT-mediated turnover. To further pursue this notion, we conducted crosslinking followed by immunoprecipitation (iCLIP) experiments37,38 with endogenous UAP56 and endogenously 3×Flag-tagged LENG8 in HeLa cells (Extended Data Fig. 8d,e and Supplementary Table 4) and quantified the iCLIP coverages of these proteins, normalized to transcript abundance, across three distinct classes of PAXT targets: pA+ ncRNAs, PROMPTs and PTTs. Although the majority of the UAP56 iCLIP signal mapped to exons (Extended Data Fig. 8f), consistent with its established role in messenger ribonucleoprotein particle (mRNP) maturation and export7,16, both UAP56 and LENG8 exhibited selective enrichment over PAXT-sensitive transcripts compared with their expression-matched and exosome-sensitive control groups (Extended Data Fig. 8g,h, Supplementary Fig. 5a,b and Methods). Thus UAP56 binds a wide range of RNAPII transcripts, including PAXT substrates. Such broad RNP incorporation presumably occurs via UAP56 recruitment through diverse set of factors, many of which include UAP56 binding motifs (UBMs)7,39,40.Fig. 4: PAXT and TREX-2 compete for pA+ RNAs.a, Box plots of RT–qPCR analysis of selected PAXT substrates as in Fig. 3b, but for cells expressing wild-type LENG8, or LENG8(F301A), LENG8(R563A) or LENG8(TRR) constructs at endogenous levels (see Extended Data Fig. 8a). b, Heat maps as in Extended Data Fig. 7c, but of pA+ RNA log2 fold abundance changes in total (left) or nuclear (Nuc) and cytoplasmic (Cyto) fractions (right) of samples of dTAGV-1-treated (4 h) cells expressing GANP–2×HA–dTAG, ZFC3H1–2×HA–dTAG or LENG8–2×HA–dTAG relative to maternal HeLa cells. Transcripts were clustered and ranked on the basis of differential expression analysis of the total GANP depletion sample. Misc., miscellaneous. c, Heat maps of mean log2 fold changes of n = 3 biological replicates of PAXT-sensitive mRNAs (DESeq2; see Fig. 3) (left) and corresponding SILAC LFQ intensity changes (right) upon dTAGV-1 treatment of cells expressing LENG8–2×HA–dTAG, ZFC3H1–2×HA–dTAG or GANP–2×HA–dTAG. mRNA–protein pairs that were significantly (adjusted P value < 0.1) differentially expressed (per DESeq2 for mRNAs and DEP for proteins) are shown. Maternal HeLa cells are shown as controls. Heat maps were sorted by descending mRNA sensitivity to ZFC3H1 depletion. LENG8, ZFC3H1 and GANP depletion-induced mRNA and SILAC abundance changes are displayed in the top three rows. d, Box plots of RT–qPCR analysis as in Fig. 3a,b, but of selected short TREX-2 or PAXT substrates as well as the LENG8 and ZFC3H1 mRNAs in nuclear and cytoplasmic fractions of biochemically fractionated maternal HeLa cells or 2×HA–dTAG-tagged cell lines subjected to dTAGV-1-induced depletion of LENG8, ZFC3H1 or GANP for 4 h. RNA fold changes were calculated relative to levels in non-treated HeLa control cells and normalized to GAPDH mRNA separately for cytoplasmic and nuclear fractions. e, Model of nuclear pA+ RNA sorting.Together, the above experiments demonstrated the targeting of UAP56-bound RNPs by PAXT–LENG8–PS in vivo, suggesting that UAP56, while being an RNA export factor, is also crucial for nuclear RNA decay. To interrogate possible relations between these two fates of UAP56-bound RNPs, we first analysed the consequence of rapid GANP depletion in cells expressing GANP–2×HA–dTAG (Extended Data Fig. 6c) by pA+ RNA-seq (Extended Data Fig. 6d). In agreement with previous studies26,41, this mainly resulted in the downregulation of short pA+ RNAs with few introns (Fig. 4b and Extended Data Fig. 8i,j), resembling PAXT-sensitive transcripts (Fig. 3d and Extended Data Fig. 6o). Indeed, GANP-sensitive RNAs were largely upregulated upon depletion of ZFC3H1 or LENG8 (Fig. 4b, left and Extended Data Fig. 8k). We therefore propose that the suppressive effect of GANP reduction reflects PAXT-mediated degradation of export-restricted transcripts. Consistently, our fractionated pA+ RNA-seq data displayed the coinciding cytoplasmic accumulation of these RNAs following LENG8 or ZFC3H1 depletion (Fig. 4b, right). Equivalent depletion experiments in HCT116 cells (Extended Data Fig. 7e,f) recapitulated these trends (Extended Data Fig. 8l). By contrast, GANP reduction did not significantly affect levels of nuclear retained multi-exonic (more than 4 exons) PAXT substrates (Extended Data Fig. 8m; note the LENG8 mRNA defying this trend, see also below).To further elaborate on an apparent competition of TREX-2 and PAXT for their target transcripts, we monitored protein production in TREX-2- or PAXT-perturbed cells. As expected, GANP depletion impaired global protein synthesis, as revealed by decreased puromycin incorporation (Extended Data Fig. 9a). Less intuitive, but consistent with prior reports on ZFC3H111,28, depletion of LENG8 or ZFC3H1 also decreased synthesis of new protein, possibly owing to overloading of ribosomes by capped ncRNAs escaping from the nucleus42. We characterized these translational alterations further through the quantitative analysis of nascent protein synthesis using stable isotope labelling by amino acids in cell culture (SILAC) followed by mass spectrometry43,44 (Supplementary Table 5). Although only a subset of proteins matching PAXT-sensitive mRNAs were detectable, depletion of LENG8 or ZFC3H1 generally increased de novo peptide synthesis, whereas GANP depletion had little or opposing effects (Fig. 4c and Extended Data Fig. 9b). We note that this occurred despite the general conditions of decreased protein synthesis (Extended Data Fig. 9a). Proteins with increased synthesis upon LENG8 or ZFC3H1 depletion were enriched for RNA-binding proteins and RNA-processing regulators (Supplementary Table 5), which included the reciprocal increases of LENG8 and ZFC3H1 themselves (Fig. 4c and Extended Data Fig. 9b, top). Although only modestly increased in the SILAC experiment (Extended Data Fig. 9b, top right), LENG8 protein upregulation upon ZFC3H1 depletion was confirmed by western blotting analysis (Extended Data Fig. 9c,d). As the ZFC3H1 mRNA similarly did not pass the DESeq2 analysis threshold (Supplementary Table 2), we used quantitative PCR with reverse transcription (RT–qPCR) to reveal that rapid depletion of LENG8 or ZFC3H1 led to the nuclear and cytoplasmic accumulation of mRNAs encoding these proteins, whereas GANP depletion decreased accumulation of LENG8, ZFC3H1 and GANP (encoded by MCM3AP) mRNA (Fig. 4d). This supports the previous notion that retained mRNAs—such as LENG8 mRNA—are subject to regulation by the PAXT pathway.In conclusion, two structurally and functionally similar modules, LENG8–PS and GANP–PS, are critical interpreters of UAP56-bound nuclear pA+ RNPs. Nucleoplasmic PAXT and NPC-associated TREX-2 utilize these equivalent biochemical modules to control nuclear pA+ RNA homeostasis by facilitating decay and export, respectively.DiscussionHere we demonstrate that short and low exon content pA+ RNAs in UAP56-bound RNPs are highly susceptible to PAXT-mediated nuclear turnover. When PAXT function is impaired, NPC-associated TREX-2 seemingly grants export to these transcripts, demonstrating that LENG8–PS and GANP–PS can target similar pA+ RNPs. Based on these findings, we propose a general model for nuclear pA+ RNA fate determination (Fig. 4e). Both TREX-2 and PAXT may engage UAP56-bound pA+ RNPs. Owing to the nucleoplasmic localization of newly made pA+ RNPs, they would first encounter PAXT. However, being an adaptor of the 3′–5′ exonucleolytic exosome, PAXT targeting only translates into efficient decay if it occurs in the vicinity of the RNA 3′ end. This condition greatly sensitizes short transcripts. By contrast, for longer RNAs, which are compacted into larger RNPs with multiple UAP56s, a PAXT encounter may merely result in the release of an UAP56 molecule. This would counteract export indirectly, however, as long as a sufficient number of UAP56 molecules remains on the transcript, nuclear export via TREX-2 is still possible. In support of this model, a recent study found that the widespread interaction of ZFC3H1 with long and multiply spliced mRNAs did not affect transcript levels upon ZFC3H1 depletion15. We note that the targeting of LENG8–PS to decay-insensitive transcripts might have a role in maintaining sufficient levels of free nuclear UAP56, without which mRNA biogenesis defects, R-loop formation and genomic instability would prevail45. The importance of such UAP56 recycling might in fact explain why budding yeast, in which the PAXT complex has been lost, still harbours the LENG8–PS homologue23. Similarly, the cytoplasmic presence of a SAC3D1-containing TREX-2-like complex in higher eukaryotes might help remove residual UAP56 from RNA after export.Although general, the proposed model may be bypassed by specific transcripts. For example, short functional pA+ mRNAs, including stress-induced transcripts, must overcome nuclear decay. How this is achieved remains an open area of research. Stress-induced mRNP reorganization might offer protection to the RNA 3′ end, or their robust transcriptional upregulation may ensure that, despite ample nucleoplasmic decay, a sufficient number of RNPs still reaches the NPC. Consistent with the latter notion, we find that PAXT-sensitive short mRNAs are expressed at higher levels than their ncRNA counterparts (Extended Data Fig. 7i). Finally, decay might also be short-circuited by gene gating, positioning a given locus in proximity to the NPC, as was recently demonstrated for MYC46,47. We also find a small subset of multi-exonic transcripts, that are sensitive to PAXT-mediated decay, violating the general nuclear pA+ RNA decay regime (Fig. 3g). We suggest that this sensitivity is enhanced by prolonged residence in the nucleus (Extended Data Fig. 7h,j), exacerbated by intron presence (Extended Data Fig. 7k). In line with nuclear retention of pre-mRNA being able to have a regulatory role33,34,35,36, our data provide evidence that the PAXT system itself is subject to such control (Fig. 4c). Thus the PAXT axis, which primarily targets short, non-functional pA+ RNAs, has been co-opted to regulate a subset of mRNAs that are sensitized either by low exon content or prolonged nuclear retention.In summary, we propose that the major opposing fates of nuclear pA+ RNA—export and decay—exploit a shared molecular logic. At its centre is RNA-bound UAP56 and its highly regulated release from the pA+ RNP. This blurs previously held categorizations of proteins as specific RNA export or decay factors, and highlight the competition for common pA+ RNP features that ensures faithful gene expression.MethodsDNA sequencesAll oligonucleotide plasmid vectors are annotated in Supplementary Table 6.Purification of UAP56 and UAP56(Δ1–43)His-tagged UAP56 constructs (10×His–3C–UAP56 or 10×His–3C–UAP56(Δ1–43), residues 44–428) were expressed in Escherichia coli BL21 DE3 RIL using autoinduction medium at 37 °C for 16 h. Following collection, cells were resuspended in lysis buffer (25 mM HEPES pH 7.9, 5% glycerol, 300 mM NaCl, 20 mM imidazole, 0.05% Tween-20, and protease inhibitors), disrupted via sonication, and clarified by centrifugation. The supernatant was sequentially filtered through 1-µm and 0.45-µm filters before affinity purification on a HisTrap HP 5 ml column (Cytiva), equilibrated in buffer A (25 mM HEPES pH 7.9, 5% glycerol, 300 mM NaCl, 20 mM imidazole). After washing with buffer A supplemented with 70 mM imidazole, bound proteins were eluted using a linear gradient of imidazole (70–200 mM in buffer A). Peak fractions were diluted in buffer B (25 mM HEPES pH 7.9, 5% glycerol, 1 mM DTT) to reduce the NaCl concentration to 100 mM and subsequently subjected to anion-exchange chromatography on a HiTrapQ 5 ml column (Cytiva), pre-equilibrated with buffer B. Elution was performed with a linear NaCl gradient (100–500 mM). Fractions containing UAP56 were concentrated and further purified via size-exclusion chromatography using a HiLoad 16/600 Superdex 200 pg column (Cytiva), equilibrated in buffer C (25 mM HEPES pH 7.9, 5% glycerol, 100 mM NaCl, 1 mM DTT). Peak fractions containing the purified protein were pooled, concentrated, flash-frozen, and stored at −80 °C.Purification of LENG8–PSM and SAC3D1–PSM
Molecular basis of polyadenylated RNA fate determination in the nucleus - Nature
Biochemical, structural and cell biological analyses reveal that UAP56 (DDX39B) assembles with a TREX-2–like module that redirects non-functional polyadenylated RNAs from export to degradation.
18,896 words~86 min read