AbstractWhale falls are biodiversity oases at seabeds1,2,3,4,5,6, yet their record from the oceans has remained sparse and fragmentary6,7. Here we report the discovery of a vast whale necropolis in the Diamantina Zone (4,616- to 7,001-m depth), extending about 1,200 km along the sea floor of the southeastern Indian Ocean. This area has a deep and extensive accumulation comprising five modern natural whale-fall communities and 476 fossil cetaceans recorded. We show that carcasses host specialized communities dominated by brittle stars, bone-boring worms and chemosynthesis-based bivalves and that the fossil record in this area comprises both extant and extinct deep-diving beaked whales. Isotopic dating shows that whale falls in this region have occurred since at least 5.3 million years ago. These findings reshape the understanding of the limits and biogeography of whale-fall ecosystems and establish some deep sea floors as a fossil archive for tracing cetacean evolution over geological time.
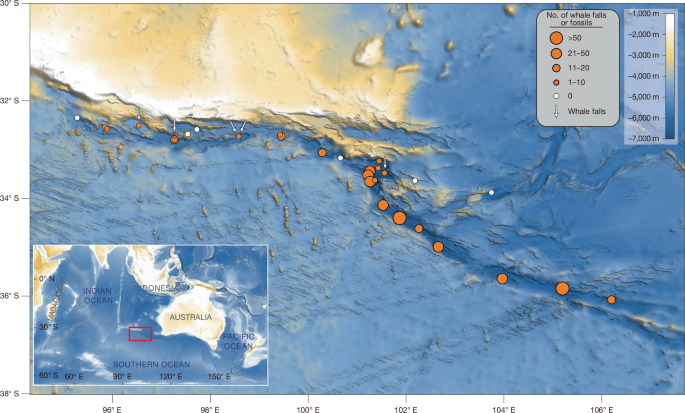
MainThe deep sea is home to myriad life forms that have adapted to extreme environmental conditions. One of the most fascinating phenomena of the deep sea are whale-fall communities, whereby a whale carcass that sinks to the ocean floor1,2,3 initiates a highly idiosyncratic ecosystem in association with a variety of physiologically diverse organisms, thus providing crucial insights into the interplay of life and death in the ocean’s depths4,5,6,7. Although whale falls are abundant, with more than 70 sites documented across diverse ocean basins and depths6, their distribution remains patchy and only sporadically documented7.The species composition and diversity of whale-fall communities are strongly influenced by water depth and related environmental factors, such as temperature and hydrodynamic regime6,8,9. In contrast to deep-sea sites, shallow-water shelf whale falls generally yield different sets of taxa, indicating that highly specialized and species-rich whale-fall communities develop primarily in the food-limited setting of the deep ocean7,10,11,12,13. So far, however, most whale falls are found between some tens of metres to around 4,000 m water depth6,7, with the deepest example reaching 4,204 m in the southwest Atlantic Ocean14. The biogeography, evolutionary novelty and connectivity of deep-sea whale-fall communities remain poorly understood, first and foremost because of the paucity of data from abyssal and hadal depths7.The Diamantina Fracture Zone lies to the south of the Broken Ridge and Perth Abyssal Plain in the Indian Ocean, stretching about 1,200 km parallel to the Southeast Indian Ridge (Fig. 1). It formed as the Australian and Antarctic continents separated between 60 million and 50 million years ago15. The rough sea floor topology is the result of block faulting16. The deepest regions of the Diamantina Zone cluster in its northwestern section, most notably at the Dordrecht Deep, which reaches a maximum depth of 7,079 m as measured by the conductivity–temperature–depth sensor aboard the human-occupied vehicle (HOV) Fendouzhe in 2023. The Diamantina Zone has not been previously documented to be associated with any whale falls.Fig. 1: Distribution and abundance of whale fossils and whale falls in the Diamantina Zone.The alternative text for this image may have been generated using AI.Full size imageOrange circles indicate dive locations where whale fossils or whale falls were observed; circle size corresponds to the number of whale remains recorded per dive. Whale falls in the sulfophilic stage are marked with white arrows. White circles denote dives with no observed whale fossil or whale fall. Notably, the distribution of both whale falls and whale fossils is restricted to the Diamantina Zone sea floor; none were detected outside this region. Figure adapted from ref. 51, Wiley, under a Creative Commons licence CC BY 4.0.Whale-fall biotaFrom 8 February to 17 March 2023, using the Fendouzhe submersible capable of reaching depths of up to 11,000 m on board the R/V Tansuoyihao, we discovered extensive whale falls and fossils in the Diamantina Zone (Fig. 1 and Supplementary Video 1). During dive FDZ159, we first encountered whale fossils at a depth of 7,002 m, near the deepest point of the Dordrecht Deep. These fossils were found partially buried in soft surface sediments and lightly coated with black Fe–Mn oxides. Following the initial discovery, we conducted 32 dives to the sea floor, aiming at mapping the spatial distribution and extent of the whale falls and fossils, as well as identifying any associated whale-fall ecosystems. By doing this, we documented and collected samples from 485 whale-fossil sites and active whale falls (Fig. 2, Extended Data Tables 1 and 2 and Supplementary Table 1) from 1,200 km along the sea floor, representing an ecological landmark in the Diamantina Zone.Fig. 2: Whale falls in the sulfophilic stage in the Diamantina Zone.The alternative text for this image may have been generated using AI.Full size imagea, A 3-m-long minke whale fall recorded during dives FDZ174/FDZ177 at 5,610 m depth. b, Close-up view of yellow box in a showing brittle stars Ophiambix sp. (white arrows) and scale worms polynoid (yellow arrows) on the cranium. c, Close-up view of white box in a showing numerous transparent tubes of the bone-eating worm Osedax sp. emergent from whale bone. d, A beaked-whale fall consisting of eight ribs and several thoracic vertebrae observed during dive FDZ178 at 5,609 m depth. Brittle stars Ophioscolecidae sp. and Silax sp. and chemosymbiotic vesicomyid bivalves Abyssogena southwardae are associated with these bones. Tubeworm Nicomache sp. occurs in the surrounding sediments. e, Close-up view of the white box in d showing three vesicomyid bivalves (white arrows) and brittle stars (yellow arrows). f, A whale bone recorded during dive FDZ173 at 4,625.2 m depth, dominated by gastropods Phymorhynchus sp. 1 (white arrows), polychaetes Nicomache sp. (yellow arrows) and Osedax sp. (orange arrows). g, The deepest whale fall, consisting of three elongated vertebrae of a beaked whale, with small gastropods (yellow arrows) on their surface, observed during dive FDZ163 at 6,788.7 m depth. The two red laser points are spaced 10 cm apart. Scale bars, 5 cm.The five active whale falls are in the sulfophilic stage (Fig. 2). Bones are covered with dense, whitish microbial mats and bone-boring worms Osedax, indicating prolonged residence on the sea floor. At 6,789 m water depth, the Dordrecht Deep beaked-whale carcass WF1, consisting of three elongated vertebrae, has the deepest active whale-fall community. The largest carcass encountered, the 5-m-long skeleton WF3, was identified as the Antarctic minke whale Balaenoptera bonaerensis through the highly diagnostic earbone morphology17 and a nearly complete mitochondrial genome (GenBank: PX519993).Across the five whale falls in the sulfophilic stage, the associated fauna are taxonomically broad, comprising 35 recognized macrofaunal taxa (more than 0.5 mm in size) documented from in situ imagery and collected specimens (Extended Data Table 1 and Extended Data Figs. 1 and 2). The macrofauna are dominated by annelids, crustaceans and molluscs, with further cnidarians and nematodes. Bone-eating worms, gastropods, vesicomyid bivalves and brittle stars dominate the megafauna (more than several centimetres in size), reaching local densities up to 2,840 individuals per square metre (Supplementary Table 2).Most recovered taxa may be new to science. Molecular data were obtained for 21 species, but only the vesicomyid bivalve Abyssogena southwardae could be confidently assigned to species level through barcoding comparison with GenBank records; all remaining species were identified at genus or family rank, integrating morphological data. Three chemosymbiotic bivalves (Adipicola sp., Abyssogena southwardae and Thyasiridae sp.) hosting different sulfur-oxidizing microbial symbionts (Extended Data Fig. 2) and two bone-eating worms (Osedax sp. 1 and Osedax sp. 2) form the core of these communities18,19. The observations at water depths of 5,609 m and 5,634 m of sea daisies (Asteroidea: Xyloplax sp.) provide, to our knowledge, the deepest evidence of this genus, as well as the first record on whale falls, expanding the habitat of Xyloplax beyond wood falls and hydrothermal vents20. Three brittle-star species (Ophiambix sp., Ophioscolecidae gen. et sp. and Silax sp.) recovered from the whale skeletons differ notably from the dominant trench-floor ophiuroid genera Ophiosphalma and Ophiuroglypha. The absence of whale-fall species in the background sediments indicates that these brittle-star assemblages are highly specialized and confined to organic-rich whale substrates. In addition, some whale falls, for instance, at depths of 5,115 m, 6,470 m, 6,570 m and 6,772 m, are in the final reef stage (Extended Data Fig. 3). The exterior of these skeletons is primarily inhabited by common hard-substrate megafauna, such as the stalked sea anemone Galatheanthemum profundale, the pedunculate sponge Caulophacus sp. and the sea star Freyastera sp. The different faunal composition of the studied whale falls may be attributable to their sites, successional stages or carcass sizes.Fossil whale faunaThe palaeontological analysis of 43 recovered fossils from the Diamantina Zone led to the identification of five beaked-whale species and one baleen-whale species. Most of the beaked-whale specimens, primarily consisting of isolated rostra, were attributed to two living ziphiid species: the Andrews’ beaked whale, Mesoplodon bowdoini (Fig. 3a,b), and the strap-toothed whale, Mesoplodon layardii (Fig. 3c,d), both of which are known to inhabit the present-day southeastern Indian Ocean21. Diagnostic traits of M. layardii preserved in 14 specimens include a narrow, elongated, transversely compressed rostrum and a strongly pachyosteosclerotic vomer forming a prominent posterior bulge, a secondary anterior bulge and a midrostral dorsal depression in between (Fig. 3c,d and Extended Data Fig. 4a–g). Further matching traits, such as the shape of the prominential notch and maxillary tubercle and the size and position of the infraorbital and premaxillary foramina, further support this identification22. The seven rostra assigned to M. bowdoini are robust, moderately elongated and laterally compressed and provided with a single anterior vomeral bulge and a ventrally deflected apex, consistent with previous descriptions22 (Fig. 3a,b and Extended Data Fig. 4e–h).Fig. 3: Fossil crania of Ziphiidae from the Diamantina Zone.The alternative text for this image may have been generated using AI.Full size imagea,b, Mesoplodon bowdoini, FDZ180-R1a dorsal (a) and lateral (b) views. c,d, Mesoplodon layardii, FDZ184-R1a dorsal (c) and lateral (d) views. e–g, Pterocetus diamantinae sp. nov., FDZ182-R1a, holotype dorsal (e), lateral (f) and anterior (g) views. h, Pterocetus benguelae, FDZ186-R3a. i. Izikoziphius rossi, FDZ163-R7a. Dotted lines indicate uncertain sutures. Oblique lines indicate principal break surfaces. Scale bar, 20 cm.Three exceptionally well-preserved skulls were identified as belonging to the extinct genera Pterocetus and Izikoziphius, which were first described from fossils trawled from the sea floor off South Africa23. As for Pterocetus, this extinct relative of the modern Mesoplodon spp. and bottlenose beaked whales is characterized by distinctive wing-like expansions of the preorbital processes23. One Pterocetus specimen from the Diamantina Zone is referred to P. benguelae (Fig. 3h); the other represents a new species, Pterocetus diamantinae sp. nov. (Supplementary Note, Fig. 3e–g, Extended Data Figs. 5a,b and 6 and Extended Data Table 3). The single Izikoziphius cranium closely resembles the type species I. rossi (Fig. 3i and Extended Data Fig. 5e,f). Izikoziphius is a close relative of the extant Cuvier’s beaked whale but is recognized as a separate genus owing to the observation of a unique fossa on the premaxilla, a dome-shaped maxillary crest and a proportionally longer rostrum23. Baleen-whale fossils include a fairly well-preserved tympanic bulla of the sei whale, Balaenoptera borealis (Extended Data Fig. 5g–j) and several dilapidated, largely indeterminate cranial and postcranial remains of mostly balaenopterid mysticetes (Extended Data Fig. 5k–l).Dating of whale fossilsTo determine the ages of the fossils, we analysed 33 fossil bone specimens for their strontium isotope composition (87Sr/86Sr) (Fig. 4 and Extended Data Table 4). This method relies on comparing the isotopic signature preserved in the biominerals to the known historical record of seawater isotopes. Although this method is typically performed on compact dental tissues, the hyperdense bones of ziphiid rostra probably preserves a pristine Sr-isotope ratio24. Eight samples exhibited Sr-isotope ratios identical to modern seawater, indicating complete geochemical exchange after death. The remaining 25 samples, however, yielded 87Sr/86Sr ratios ranging from 0.709173 to 0.709029. When calibrated against the seawater 87Sr/86Sr curve25, these values correspond to ages between 0.12 Ma and 5.26 Ma. The fossil species Pterocetus bengulae and Izikoziphius rossi were found to be the oldest, with Sr-isotope average ages of 5.26 Ma and 2.44 Ma, respectively, whereas the extant species M. bowdoini (1.14–0 Ma) and M. layardii (1.20–0 Ma) are geologically younger. The oldest date indicates that whale-fall events have occurred in the Diamantina Zone since at least the Early Pliocene.Fig. 4: Strontium isotope dating of whale fossils from the Diamantina Zone.The alternative text for this image may have been generated using AI.Full size imageThe 87Sr/86Sr ratios are shown for a total of 33 samples, comprising 21 beaked whales (Ziphiidae), five baleen whales and seven unidentified cetacean specimens. Error bars indicate the range from minimum to maximum age (vertical) and plus or minus two standard errors (horizontal). Details in Extended Data Table 4.Genesis of the whale necropolisOn the basis of submersible observations, the density of whale remains reaches up to 759.5 individuals per square kilometre. The concentration of whale falls and fossils in the Diamantina Zone raises fundamental questions about the origin of this whale necropolis. Active whale-fall ecosystems were found thriving around both baleen and beaked-whale carcasses. The former group also includes the skeleton of an Antarctic minke whale, a circumpolar migratory species that is known to travel northward into the waters off southern Australia26,27. This epipelagic filter-feeder relies on krill in the upper ocean layers, mostly at depths not greater than 150 m (ref. 28). Most cetacean fossils consist of robust beaked-whale rostra, which have probably endured the destructive biostratinomic processes at play on the sea floor thanks to their hyperostotic structure. Among the few exceptions are poorly diagnostic bone fragments of baleen-whale skulls and the well-mineralized tympanic bulla of a sei whale. Like the Antarctic minke whale, the sei whale migrates seasonally into the southeastern Indian Ocean29. Observations of the diving behaviour of sei whales indicate feeding on copepods to depths of 50 m (ref. 30). Thus, the occurrence of remains of B. bonaerensis and B. borealis at such hadal depths is not related to deep-diving habits and rather is due to the carcasses sinking to the sea floor of this shared migratory corridor.The vast majority of the cetacean remains belong to two deep-diving ziphiid species: the strap-toothed and Andrews’ beaked whales, both of which are known to inhabit the southeastern Indian Ocean21,31,32. Beaked whales are specialized predators of deep-water squid and fish, foraging along steep continental slopes, submarine canyons, abyssal plains and trenches33. The Diamantina Zone, with its extreme depths ranging between 4,200 m and 7,000 m, complex V-shaped topography and abundant squid and fish resources as observed during our dives, provides an ideal deep-water foraging ground for beaked whales.Natural mortality, combined with the inherent risks of deep diving, probably contributes to the accumulation of beaked-whale remains in the sea floor of this zone. These beaked whales possess extraordinary physiological adaptations for deep diving, routinely reaching depths more than 1,000 m and holding their breath for more than a hour33,34,35. The maximum dive depth for beaked whales is estimated to be more than 3,000 m on the basis of lung collapse and oxygen storage34,35,36,37,38. Thus, foraging at depths exceeding 3,000 m would be too physiologically taxing for beaked whales and may heighten the risk of fatal exhaustion or decompression sickness37,38,39. Ultimately, the V-shaped topography of the Diamantina Zone may further contribute to this accumulation by funnelling and concentrating onto the sea floor the sinking carcasses caused by natural and accidental mortality.Critically, the ultra-low regional sedimentation rate close to the Diamantina Zone (Broken Ridge, 0–5 Ma, 0.05–0.55 cm kyr−1)40 implies a prolonged exposure of the skeletal remains at the sea floor: one that would probably last more than several hundred thousand years at least. On slopes or uplifted sea floor zones, skeletal remains may remain exposed for extended periods: up to 5.3 million years, according to our dating data. The fossilized remains we observed are almost exclusively beaked-whale rostra, some of which have the highest bone density and mineral content among extant vertebrates41. This high compactness probably inhibits rapid degradation, with long-term preservation on the sea floor being further enhanced by the progressive accumulation of ferromanganese oxides both within the bone matrix and on the outer bone surface. The latter process isolates the skeletal elements from the surrounding environment while increasing their robustness. For buried bones, additionally, authigenic carbonate precipitation during the organic degradation may also facilitate fossil preservation. The confluence of beaked whales’ deep-diving ecology, extreme foraging physiology, topographic focusing, an ultra-low sedimentation rate and early fossilization may explain the formation of this whale necropolis.ImplicationsThe discovery of whale-fall communities in the Diamantina Zone at depths exceeding 6,700 m establishes one of the deepest known whale-fall ecosystems in the ocean, extending the known depth range of such habitats by more than 2,500 m. Isolation, imposed by extreme depth, apparently has facilitated the development of a distinct, specialized whale-fall community dominated by species that may be new to science, as indicated by our molecular data (Extended Data Table 1). This not only expands our understanding of metazoan species richness in the deep-sea ecosystems but, given that we are still in the early stages of discovery of deep-sea whale-fall fauna7, also indicates that these species probably exhibit ecological novelty and represent cases of adaptive radiation. For instance, these whale falls share key ecological and evolutionary links with deep-sea cold seeps and hydrothermal vents6,19, including those in hadal trenches42,43, as evidenced by shared taxa such as chemosymbiotic bivalves (Adipicola, Abyssogena and Thyasiridae), gastropods (Phymorhynchus) and squat lobsters (Munidopsis). The results support the hypothesis that deep-sea whale falls act as evolutionary hotspots and biogeographic stepping stones for sulfide-dependent fauna in the deep ocean2,7,44,45,46,47. In the total survey area of 0.64 km2 (from 32 dives), five active whale falls were observed. This yields a density of 7.81 whale falls per square kilometre. Aligned along a northwest–southeast axis for 1,200 km, these falls may form a previously unrecognized ‘whale-fall community supercorridor’. This extensive biogeographic feature could have an important role in the dispersal, connectivity and evolution of deep-sea chemosynthetic communities across the Southern Indian Ocean.As beaked whales are known primarily from rare strandings, their abundance, distribution and ecology remain poorly understood overall48. Our discovery of an accumulation of skeletal remains of the two extant beaked-whale species Mesoplodon bowdoini and M. layardii provides an unparalleled source of information on these largely enigmatic cetaceans. Moreover, the investigated whale fossils, preserved for more than 5 million years, serve as an archive providing a direct, continuous record for tracing evolutionary trajectories. Comparative anatomical analysis of these remains can elucidate feeding behaviours, locomotion and ecological roles of deep-diving cetaceans. Thus, the Diamantina Zone necropolis constitutes a deep-sea fossil megasite: one that offers a window into the evolutionary history, palaeoecology and population dynamics of beaked whales from the Pliocene to the present day. Similar whale necropolises probably exist in other core beaked-whale habitats, such as South Africa23, the Iberian Peninsula49 and off the Crozet and Kerguelen islands50, as indicated by the recovery of abundant fossils by trawling, indicating that comparable hidden archives may be widespread in the global deep oceans.Systematic palaeontology












